Chapter: Clinical Dermatology: The function and structure of the skin
Molecular components of the skin immune system
Molecular
components of the skin immune system
Antigens and haptens
Antigens
are molecules that are recognized by the immune system thereby provoking an
immune reac-tion, usually in the form of a humoral or cell-bound antibody
response. Haptens, often chemicals of low molecular weight, cannot provoke an
immune reac-tion themselves unless they combine with a protein. They are
important sensitizers in allergic contact dermatitis.
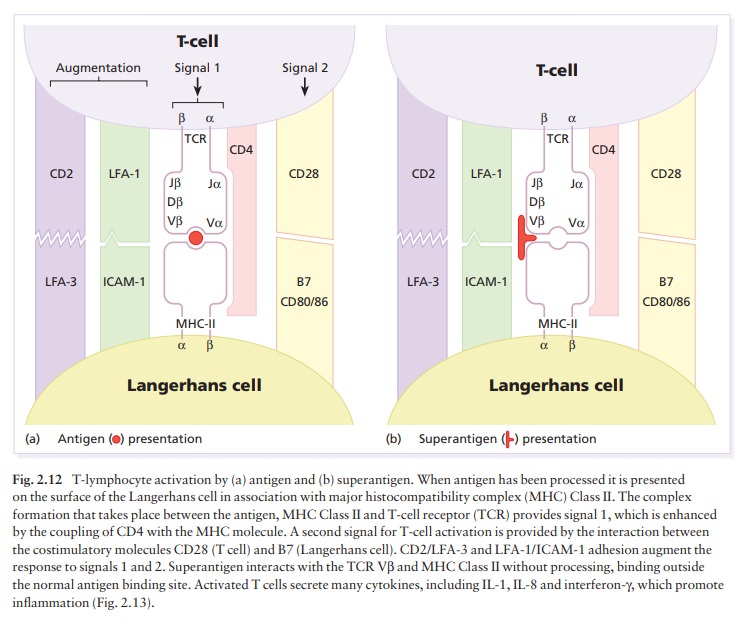
Superantigens
Some
bacterial toxins (e.g. those released by Staphylococcus aureus)
are prototypic superantigens.Sensitization to such superantigens is not
necessary to prime the immune response. Superantigens align with a variety of
MHC Class II molecules outside their antigen presentation groove and, without
any cellular processing, may directly signal to different classes of T cells
within the large family carrying a Vβ
type of T-cell receptor (Fig. 2.12). By these means, super-antigens can induce
massive T-cell proliferation and cytokine production leading to disorders such
as the toxic shock syndrome. Streptococcal toxins act as superantigens to
activate T cells in the pathogenesis of guttate psoriasis.
Antibodies (immunoglobulins)
Immunoglobulin
G (IgG) is responsible for most of the secondary response to most antigens. It
can cross the placenta, and binds complement to activate the classical
complement pathway. IgG can coat neutro-phils and macrophages (by their FcIgG
receptors), and acts as an opsonin by cross-bridging antigen. IgG can also
sensitize target cells for destruction by K cells. IgM is the largest
immunoglobulin molecule. It is responsible for much of the primary response
and, like IgG, it can fix complement but it cannot cross the placenta. IgA is
the most common immunoglobulin in secretions. It does not bind complement but
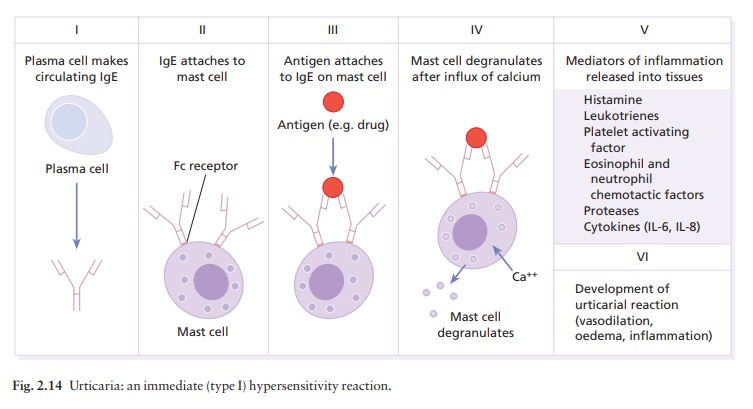
can activate complement via the alternative pathway. IgE binds to Fc receptors
on mast cells and basophils, where it sensitizes them to release inflammatory
medi-ators in type I immediate hypersensitivity reactions (Fig. 2.14).
Cytokines
Cytokines
are small proteins secreted by cells such as lymphocytes and macrophages, and
also by ker-atinocytes (Table 2.2). They regulate the amplitude and duration of
inflammation by acting locally on nearby cells (paracrine action), on those
cells that secreted them (autocrine) and occasionally on distant target cells
(endocrine) via the circulation. The term cytokine covers interleukins,
interferons, colony-stimulating factors, cytotoxins and growth factors.
Interleukins (IL) are produced predominantly by leucocytes, have a known amino
acid sequence and are active in inflammation or immunity.
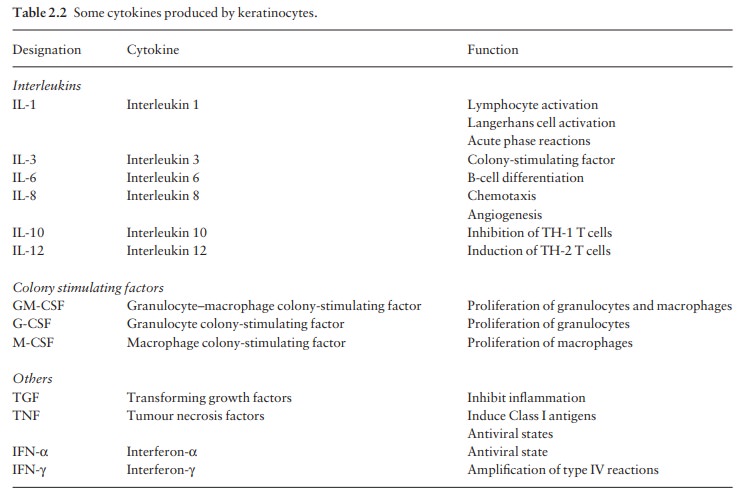
There
are many cytokines (Table 2.2), and each may act on more than one type of cell
causing many different effects. Cytokines frequently have overlapping actions.
In any inflammatory reaction some cytokines
This network
of potent chemicals, each acting alone and in concert, moves the inflammatory
response along in a controlled way. Cytokines bind to high affinity (but not
usually specific) cell surface re-ceptors, and elicit a biological response by
regulating the transcription of genes in the target cell via signal
transduction pathways involving, for example, the Janus protein tyrosine kinase
or calcium influx systems. The biological response is a balance between the
pro-duction of the cytokine, the expression of its receptors on the target
cells, and the presence of inhibitors.
Adhesion molecules
Cellular
adhesion molecules (CAMs) are surface gly-coproteins that are expressed on many
different types of cell; they are involved in cell–cell and cell–matrix
adhesion and interactions. CAMs are fundamental in the interaction of
lymphocytes with antigen-presenting cells (Fig. 2.12), keratinocytes and
endothelial cells and are important in lymphocyte trafficking in the skin
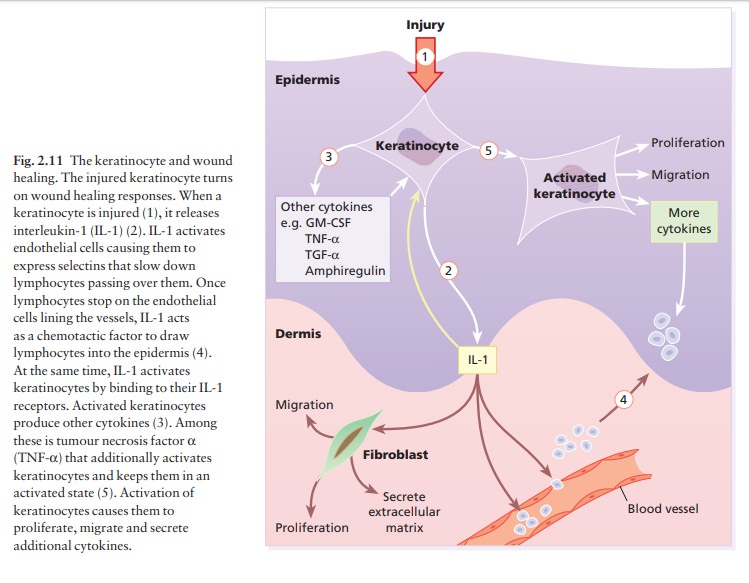
during inflammation (Fig. 2.11). CAMs have been classified into four families:
cadherins, immunoglobulin superfamily, integrins and selectins. E-cadherins are
found on the surface of keratinocytes between the desmosomes. γ-Interferon causes up-regulation of Fas on epidermal
lymphocytes. Interaction of these with Fas ligand on keratinocytes causes
e-cadherins to ‘disappear’ leading to intercellular oedema (spongiosis) between
desmosomes.
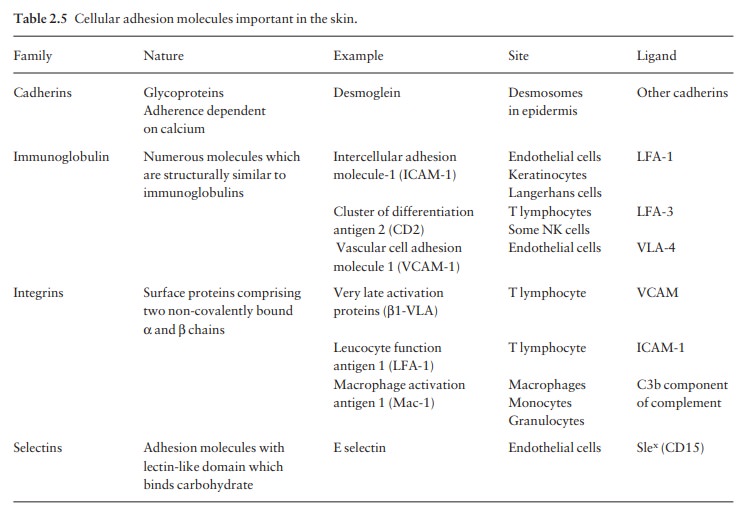
CAMs
of special relevance in the skin are listed in Table 2.5.
Histocompatibility antigens
Like
other cells, those in the skin express surface antigens directed by genes of
the MHC. The human leucocyte antigen (HLA) region lies on chromosome 6. In
particular, HLA-A, -B and -C antigens (the Class I antigens) are expressed on
all nucleated cells includ-ing keratinocytes, Langerhans cells and cells of the
dermis. HLA-DR, -DP, -DQ and -DZ antigens (the Class II antigens) are expressed
only on some cells (e.g. Langerhans cells). They are poorly expressed on
keratinocytes except during certain reactions (e.g. allergic contact
dermatitis) or diseases (e.g. lichen planus). Helper T cells recognize antigens
only in the presence of cells bearing Class II antigens. Class II antigens are
also important for certain cell–cell inter-actions. On the other hand, Class I
antigens mark target cells for cell-mediated cytotoxic reactions, such as the
rejection of skin allografts and the destruction of cells infected by viruses.
Related Topics