Chapter: Essential Microbiology: Antimicrobial Agents
Microbial associations with plants
Microbial
associations with plants
The roots of almost all plants form mutualistic
associations with fungi, known as mycorrhizae,
which serve to enhance the uptake of water and mineral nutrients, especially
phosphate, by the plants. The beneficial effect of a mycorrhizal association is
particularly noticeable in soils with a poor phosphorus content. In return, the
plant supplies reduced carbon in the form of carbohy-drates to the fungi.
Unlike other plant–microorganism interactions that occur in the rhizosphere,
mycor-rhizal associations involve the formation of a distinct,
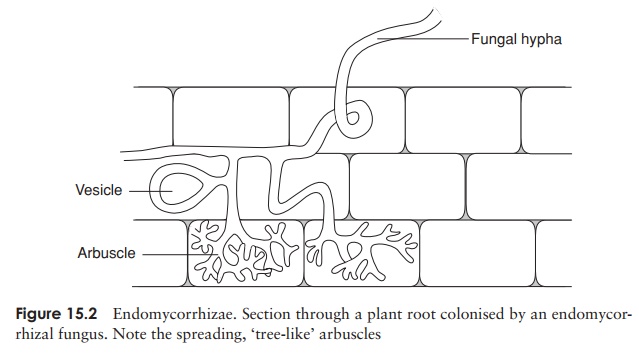
integrated structure comprising root cells and fungal
hyphae. In ectomycorrhizae the plant
partner is always a tree; the fungus surrounds the root tip, and hyphae spread
between (but do not enter) root cells. In the case of the more common endomycor-rhizae, the fungal hyphae
actually penetrate the cells by releasing cellulolytic enzymes.Arbuscular
mycorrhizae are found in practically all plant types, including ‘lower’ plants
(mosses, ferns). They form highly branched arbuscules
within the root cells that gradu-ally lyse, releasing nutrients into the plant
cells (Figure 15.2). In contrast to pathogenic fungi, mycorrhizal fungi are
often rather non-specific in their choice of ‘partner’ plant.
An interesting example of mutualistic association
concerns the endophytic (=‘inside plant’) fungus Acremonium.
The ability of crop plants to thrive is frequently
limited by the supply of available nitrogen; although there is a lot of it in
the atmosphere, plants are unable to utilise it, and instead must rely on an
inorganic supply (both naturally-occurring and in the form of fertilisers). As
we saw, however, certain bacterial species are able to ‘fix’ atmospheric
nitrogen into a usable form. Some of these, notably Rhizobium spp. form a mutualistic relationship with leguminous
plants such as peas, beans and clover, converting nitrogen to ammonia, which
the legume can incorporate into amino acids. In return, the bacteria receive a
supply of organic carbon, which they can use as an energy source for the
fixation of nitrogen.
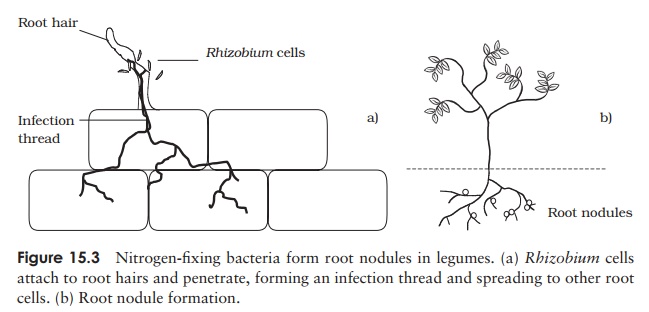
The free-living Rhizobium
enters the plant via its root hairs, forming an infection thread and infecting
more and more cells (Figure 15.3). Normally rod-shaped, they proliferate as
irregularly-shaped bacteroids,
densely packing the cells and causing them to swell, forming rootnodules.
Rhizobium requires oxygen
as a terminal electron acceptor in oxidative phosphory-lation, but as you may
recall, the nitrogenase enzyme, which fixes the nitrogen, is sensitive to
oxygen. The right microaerophilic conditions are maintained
by means of a unique oxygen-binding pigment, leghaemoglobin. This is only
synthe-sised by means of a collaboration between both partners. Nitrogen
fixation requires a considerable input of energy in the form of ATP (16
molecules for every molecule of nitrogen), so when ammonia is in plentiful
supply the synthesis of the nitrogenase enzyme is repressed.
Farmers have long recognised the value of
incorporating a legume into a crop rotation system; the nodules left behind in
the soil after harvesting the crop appreciably enhance the nitrogen content of
the soil.
Legumes are not the only plants able to benefit from
the nitrogen-fixing capabilities of bacteria. The water fern Azolla, which grows prolifically in the
paddy fields of southeast
Asia, has its nitrogen supplied by the blue-green bacterium Anabaena.
When the fern dies, it acts as a natu-ral fertiliser for the rice crop. Anabaena does not form root nodules, but
takes up residence in small pores in the Azolla
fronds. Nitrogen fixation takes place in hetero-cysts, specialised cells
whose thick walls slow down the rate at which oxygen can diffuse into the cell,
providing appropriate conditions for the oxygen-sensitive nitrogenase.
The alder tree (Alnus
spp.) is able to grow in soils with poor nitrogen content due to its
association in root nodules with the nitrogen-fixing actinomycete Frankia. The filamentous Frankia solves the problem of
nitrogenase’s sensitivity to oxygen by com-partmentalising it in thick-walled
vesicles at the tips of its hyphae, which serve the same function as the
heterocysts of Anabaena.
Many microorganisms, particularly bacteria and
yeasts, are to be found living as harmless commensals on the surface structures
(leaves, stem, fruits) of plants.
Plant disease may be caused by viruses, bacteria,
fungi or protozoans. These frequently have an impact on hu-mans, especially if
the plant affected is a commercially important crop. Occasionally the effecton
a human

population can be catastrophic, as with the Irish
famine of the 1840s brought about by potato blight. A number of microbial
diseases of plants are listed in Table 15.5We have already encountered the soil
bacterium Agrobacterium tumefaciens
in Chap-ter 12, where we saw how it has been exploited as a means of
genetically modifying plants. A.
tumefaciens is useful for introducing foreign DNA because it is a natural
pathogen of plants, entering wounds and causing crown gall disease, a condition
char-acterised by areas of uncontrolled growth, analogous to tumour formation
in animals. This proliferation is caused by the expression within the plant
cell of genes that en-code the sequence for enzymes involved in the synthesis
of certain plant hormones. The genes are carried on the T-DNA, part of an A. tumefaciens plasmid, which integrates
into a host chromosome. Also on the T-DNA are genes that code for amino acids
called opines. These are of no value to the plant, but are utilised by the A. tumefaciens as a food source.


Related Topics