Chapter: Medical Physiology: Membrane Physiology, Nerve, and Muscle : Contraction of Skeletal Muscle
Mechanics of Skeletal Muscle Contraction
Mechanics of Skeletal Muscle Contraction
Motor Unit. Each motoneuron that leaves the spinal cordinnervates multiple muscle fibers, the number depend-ing on the type of muscle. All the muscle fibers inner-vated by a single nerve fiber are called a motor unit. In general, small muscles that react rapidly and whose control must be exact have more nerve fibers for fewer muscle fibers (for instance, as few as two or three muscle fibers per motor unit in some of the laryngeal muscles). Conversely, large muscles that do not require fine control, such as the soleus muscle, may have several hundred muscle fibers in a motor unit. An average figure for all the muscles of the body is questionable, but a good guess would be about 80 to 100 muscle fibers to a motor unit.
The muscle fibers in each motor unit are not all bunched together in the muscle but overlap other motor units in microbundles of 3 to 15 fibers. This interdigita-tion allows the separate motor units to contract in support of one another rather than entirely as individ-ual segments.
Muscle Contractions of Different Force – Force Summation.
Summation means the adding together of individualtwitch contractions to increase the intensity of overall muscle contraction. Summation occurs in two ways: (1) by increasing the number of motor units contracting simultaneously, which is called multiple fiber summa-tion, and (2) by increasing the frequency of contraction,which is called frequency summation and can lead to tetanization.
Multiple Fiber Summation. When the central nervoussystem sends a weak signal to contract a muscle, the smaller motor units of the muscle may be stimulated in preference to the larger motor units. Then, as the strength of the signal increases, larger and larger motor units begin to be excited as well, with the largest motor units often having as much as 50 times the contractile force of the smallest units. This is called the size princi-ple. It is important, because it allows the gradations ofmuscle force during weak contraction to occur in small steps, whereas the steps become progressively greater when large amounts of force are required. The cause of this size principle is that the smaller motor units are driven by small motor nerve fibers, and the small motoneurons in the spinal cord are more excitable than the larger ones, so they naturally are excited first.
Another important feature of multiple fiber summa-tion is that the different motor units are driven asyn-chronously by the spinal cord, so that contraction alternates among motor units one after the other, thus providing smooth contraction even at low frequencies of nerve signals.
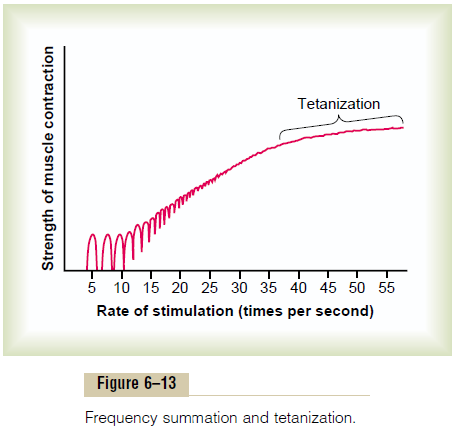
Frequency Summation and Tetanization. Figure 6–13shows the principles of frequency summation and tetanization. To the left are displayed individual twitch contractions occurring one after another at low fre-quency of stimulation. Then, as the frequency increases, there comes a point where each new contraction occurs before the preceding one is over. As a result, the second contraction is added partially to the first, so that the total strength of contraction rises progressively with increasing frequency. When the frequency reaches a critical level, the successive contractions eventually become so rapid that they fuse together, and the whole muscle contraction appears to be completely smooth and continuous, as shown in the figure. This is called tetanization. At a slightly higher frequency, the strengthof contraction reaches its maximum, so that any addi-tional increase in frequency beyond that point has no further effect in increasing contractile force. This occurs because enough calcium ions are maintained in the muscle sarcoplasm, even between action potentials, so that full contractile state is sustained without allowing any relaxation between the action potentials.
Maximum Strength of Contraction. The maximum strengthof tetanic contraction of a muscle operating at a normal muscle length averages between 3 and 4 kilograms per square centimeter of muscle, or 50 pounds per square inch. Because a quadriceps muscle can have as much as 16 square inches of muscle belly, as much as 800 pounds of tension may be applied to the patellar tendon. Thus, one can readily understand how it is possible for muscles to pull their tendons out of their insertions in bone.
Changes in Muscle Strength at the Onset of Contraction—The Staircase Effect (Treppe). When a muscle begins to con-tract after a long period of rest, its initial strength of contraction may be as little as one half its strength 10 to 50 muscle twitches later. That is, the strength of con-traction increases to a plateau, a phenomenon called the staircase effect, or treppe.
Although all the possible causes of the staircase effect are not known, it is believed to be caused primarily by increasing calcium ions in the cytosol because of the release of more and more ions from the sarcoplasmic reticulum with each successive muscle action potential and failure of the sarcoplasm to recapture the ions immediately.
Skeletal Muscle Tone. Even when muscles are at rest, acertain amount of tautness usually remains. This is called muscle tone.Because normal skeletal muscle fibers do not contract without an action potential to stimulate the fibers, skeletal muscle tone results entirely from a low rate of nerve impulses coming from the spinal cord. These, in turn, are controlled partly by signals transmitted from the brain to the appropriate spinal cord anterior motoneurons and partly by signals that originate in muscle spindles located in the muscle itself. Both of these are discussed in relation to muscle spindle and spinal cord function.
Muscle Fatigue. Prolonged and strong contraction of amuscle leads to the well-known state of muscle fatigue. Studies in athletes have shown that muscle fatigue increases in almost direct proportion to the rate of depletion of muscle glycogen. Therefore, fatigue results mainly from inability of the contractile and metabolic processes of the muscle fibers to continue supplying the same work output. However, experiments have also shown that transmission of the nerve signal through the neuromuscular junction, can diminish at least a small amount after intense pro-longed muscle activity, thus further diminishing muscle contraction. Interruption of blood flow through a con-tracting muscle leads to almost complete muscle fatigue within 1 or 2 minutes because of the loss of nutrient supply, especially loss of oxygen.
Lever Systems of the Body. Muscles operate by applyingtension to their points of insertion into bones, and the bones in turn form various types of lever systems. Figure 6–14 shows the lever system activated by the biceps muscle to lift the forearm. If we assume that a large
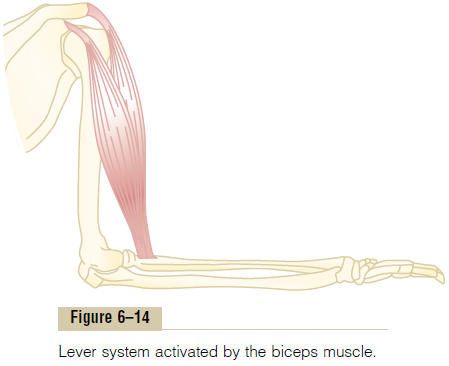
biceps muscle has a cross-sectional area of 6 square inches, the maximum force of contraction would be about 300 pounds. When the forearm is at right angles with the upper arm, the tendon attachment of the biceps is about 2 inches anterior to the fulcrum at the elbow, and the total length of the forearm lever is about 14 inches. Therefore, the amount of lifting power of the biceps at the hand would be only one seventh of the 300 pounds of muscle force, or about 43 pounds. When the arm is fully extended, the attachment of the biceps is much less than 2 inches anterior to the fulcrum, and the force with which the hand can be brought forward is also much less than 43 pounds.
In short, an analysis of the lever systems of the body depends on knowledge of (1) the point of muscle inser-tion, (2) its distance from the fulcrum of the lever, (3) the length of the lever arm, and (4) the position of the lever. Many types of movement are required in the body, some of which need great strength and others of which need large distances of movement. For this reason, there are many different types of muscle; some are long and contract a long distance, and some are short but have large cross-sectional areas and can provide extreme strength of contraction over short dis-tances. The study of different types of muscles, lever systems, and their movements is called kinesiology and is an important scientific component of human physioanatomy.
“Positioning” of a Body Part by Contraction of Agonist and Antag-onist Muscles on Opposite Sides of a Joint—“Coactivation” of Antagonist Muscles. Virtually all body movements arecaused by simultaneous contraction of agonist and antagonist muscles on opposite sides of joints. This is called coactivation of the agonist and antagonist muscles, and it is controlled by the motor control centers of the brain and spinal cord.
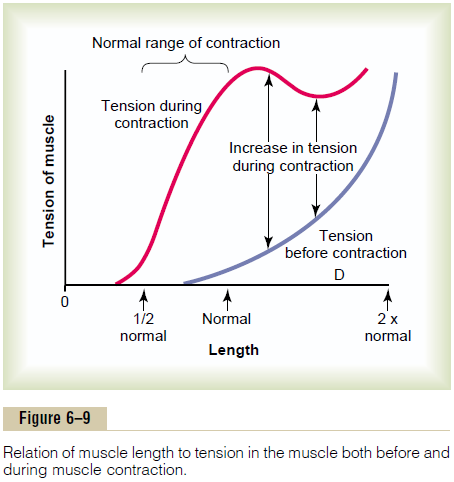
The position of each separate part of the body, such as an arm or a leg, is determined by the relative degrees of contraction of the agonist and antagonist sets of muscles. For instance, let us assume that an arm or a leg is to be placed in a midrange position. To achieve this, agonist and antagonist muscles are excited about equally. Remember that an elongated muscle contracts with more force than a shortened muscle, which was demonstrated in Figure 6–9, showing maximum strength of contraction at full functional muscle length and almost no strength of contraction at half normal length. Therefore, the elongated muscle on one side of a joint can contract with far greater force than the shorter muscle on the opposite side. As an arm or leg moves toward its midposition, the strength of the longer muscle decreases, whereas the strength of the shorter muscle increases until the two strengths equal each other. At this point, movement of the arm or leg stops. Thus, by varying the ratios of the degree of activation of the agonist and antagonist muscles, the nervous system directs the positioning of the arm or leg.
Related Topics