Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Zoogeography
Marine zoogeographic regions fishes
Marine zoogeographic regions
Global biogeographic patterns in the marine environment were outlined by Ekman (1953, but first published in German in 1935) and Briggs (1974, 1995). Briggs’ patterns focused on a system of coastal and shelf provinces partly defined by their degree of endemism. A system of 64 large marine ecosystems (LMEs) was developed over recent years by Kenneth Sherman and others. LMEs are relatively large areas of 200,000 km2 or greater characterized by distinct bathymetry, hydrography, productivity, and trophically dependent populations (Sherman et al. 2005). About 90% of the world’s annual yield of marine fisheries is produced within the boundaries of the 64 LMEs. A new global system for coastal and shelf areas, Marine Ecosystems of the World (MEOW), is a hierarchical nested system of 12 realms containing 62 provinces and 232 ecoregions (Spalding et al. 2007).
The simplest division of the distributions of inshore marine fishes is into four major marine regions, in order of decreasing biodiversity: (i) Indo-West Pacific, (ii) western Atlantic, (iii) eastern Pacific, and (iv) eastern Atlantic. These regions are separated from each other either by continents or by large expanses of open ocean, and each has been subdivided into different units by different authors.
Indo-West Pacific region
The Indo-West Pacific region – from South Africa and the Red Sea east through Indonesia and Australia to Hawaii and the South Pacific Islands, all the way to Easter Island – contains about one-third of the species of shallow marine fishes, about 3000 species, compared to no more than 1200 in any other region. Multiple datasets show global maxima of marine biodiversity to be in the Indo-Malay-Philippines Archipelago. Analysis of distribution data for 2983 marine species of fishes, other vertebrates, and invertebrates reveals a pattern of richness on a finer scale and identifi es a peak of marine biodiversity in the central Philippine Islands and a secondary peak between peninsular Malaya and Sumatra (Carpenter & Springer 2005). Biodiversity is also high in many other marine taxa in this region (Briggs 1974). There are approximately 70 genera and about 500 species of hermatypic (reef-building) corals in this region, which is 10 times the number of species present in the western Atlantic (Rosen 1988). Among other groups, the Indo-West Pacific contains about 1000 species of bivalve mollusks (including all the giant clams, Tridacnidae), twice that in the western Atlantic; and 49 of the 50 species of sea snakes (Hydrophiidae), compared to one species in the eastern Pacific. Some families of fishes such as the whitings (Sillaginidae) and rabbitfishes (Siganidae) are endemic to the Indo-West Pacific.
The Pacific Plate (Fig. 16.3) forms a major biogeographic unit of the Indo-West Pacific. The Pacific Plate is the largest of the earth’s lithospheric plates and occupies most of the area that has been referred to as the Pacific Basin (Springer 1982). The number of taxa decreases sharply as one proceeds eastward across the western margin of the plate. In addition, there is a high degree of endemism on the plate.
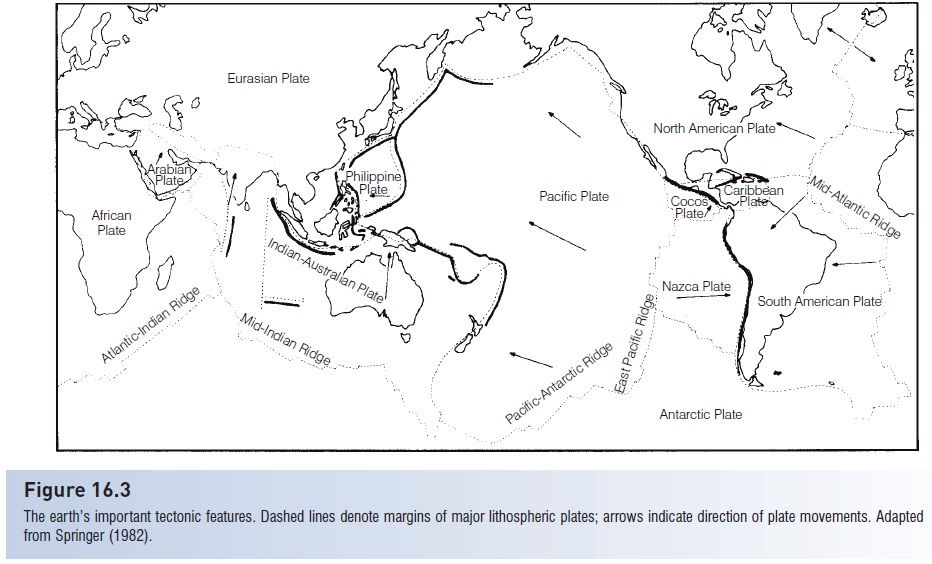
Figure 16.3
The earth’s important tectonic features. Dashed lines denote margins of major lithospheric plates; arrows indicate direction of plate movements. Adapted from Springer (1982).
An instructive example of the kinds of distributions one finds associated with specific regions and plates occurs with Spanish mackerels of the genus Scomberomorus(Collette & Russo 1985b). Of the 18 species in the genus, 10 occur in the Indo-West Pacific, but they are noticeably absent from the Pacific Plate (Springer 1982, fi g. 40). One species, Scomberomorus commerson, is widespread throughout much of the Indo-West Pacific. This distributional pattern cannot tell us much because, as with plesiomorphic characters, widespread species are not as informative about the causes of the distribution patterns as those with more restricted distributions.
In contrast, the ranges of three species, S. guttatus, S. koreanus, and S. lineolatus (Fig. 16.4), stop at the continental margin, at what is known as Wallace’s Line (see below under Oriental region). Australia and southern New Guinea, east of Wallace’s Line, have a Spanish mackerel fauna that consists of four different species: S. multiradiatus,S. semifasciatus, S. queenslandicus, and S. munroi. These four species do not extend into the East Indies or even to the north coast of New Guinea, although they easily could swim that far. This distribution pattern is obviously not simply a result of present ecological factors but instead must be historical, related to the earlier evolution and dispersal of the genus. The present island of New Guinea resulted from the collision of two plates that may have contained two different fish faunas.
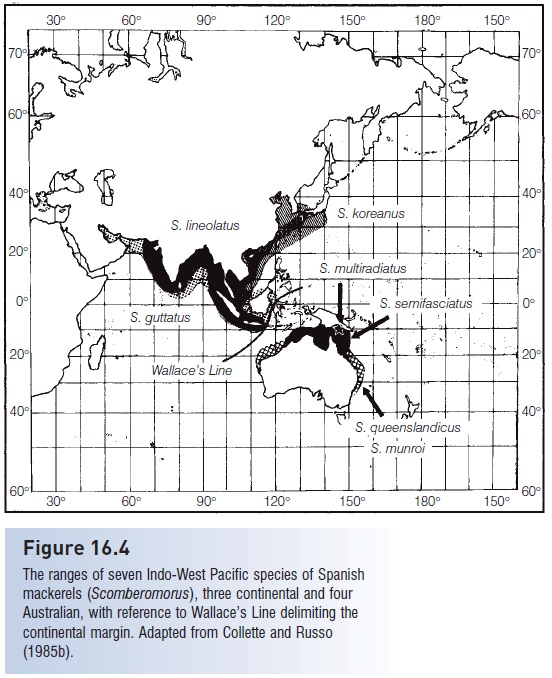
Figure 16.4
The ranges of seven Indo-West Pacific species of Spanish mackerels (Scomberomorus), three continental and four Australian, with reference to Wallace’s Line delimiting the continental margin. Adapted from Collette and Russo (1985b).
Western Atlantic region
The western Atlantic region includes the temperate shores of North America, the Gulf of Mexico, the tropical shores of the Caribbean Sea, and the tropical and temperate shores of South America. Genetic data indicate that two islands, Ascension and St. Helena, once thought to be part of the eastern Atlantic actually show more similarities to Brazil in the western Atlantic.
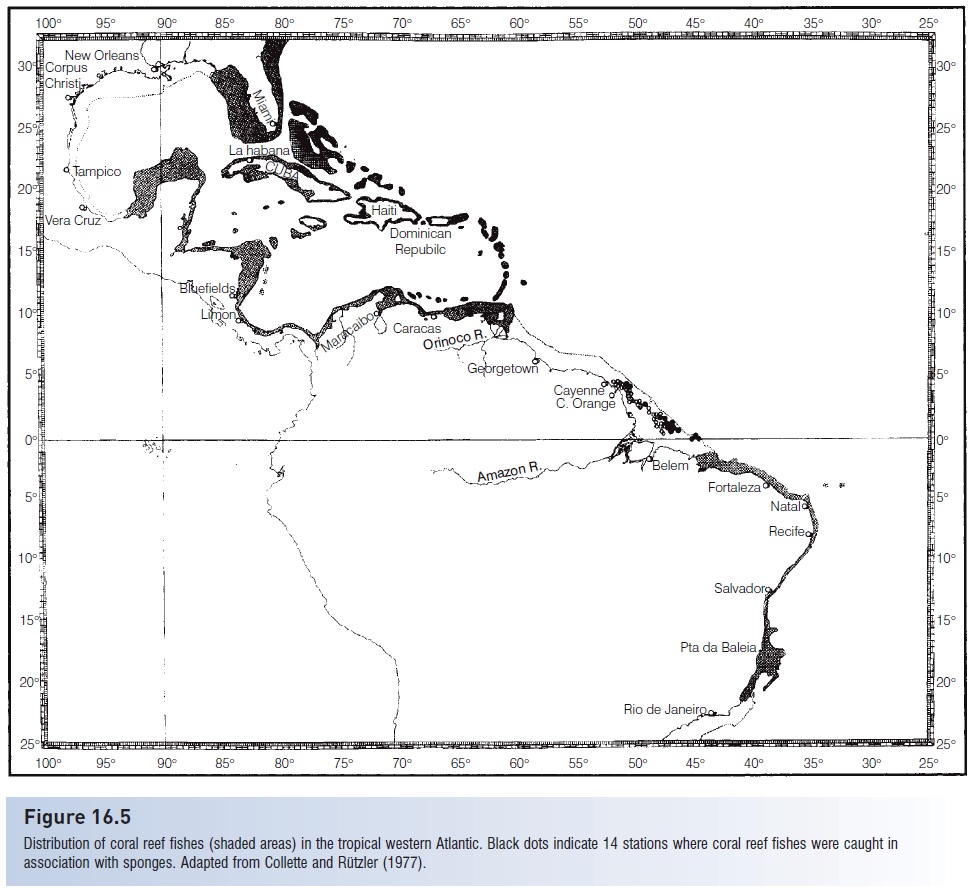
Figure 16.5
Distribution of coral reef fishes (shaded areas) in the tropical western Atlantic. Black dots indicate 14 stations where coral reef fishes were caught in association with sponges. Adapted from Collette and Rützler (1977).
The radiation of tropical shore fishes in the western Atlantic is associated with the presence of the habitat created by West Indian coral reefs; 16–24 genera of zooxanthellate (containing symbiotic algae) corals occur there (Rosen 1988). The fish fauna of the western Atlantic region comprises about 1200 species. This coral reef fish fauna is partially divided into northern and southern parts by the freshwater outflow of the Amazon River. Bottom trawling below the freshwater outflow of the Amazon River in 1975 closed part of the supposed “gap” in fish distributions (Collette & Rützler 1977). At 14 benthic stations off the mouth of the Amazon under the superfi cial freshwater layer, a typical reef fish fauna of 45 species was found, but these “coral reef ” species were associated with 35 species of sponges in water too turbid for coral growth (Fig. 16.5).
Sponges provide the necessary structural habitat for “coral reef ” species, which allows genetic continuity between the two supposedly separated populations. However, shallower water species are separated by the fresh waters of the Amazon outflow (Floeter et al. 2008). Rocha et al. (2002) showed the variable effects of the Amazon outflow on three species of the surgeonfish genus Acanthurus. The Amazon outflow is a strong barrier to dispersal of A. bahianus, a modest barrier for A. coeruleus, and has no discernable effect on A. chirurgus, which has been collected on deep soft bottoms with sponge habitats under the Amazon outflow. Both A. bahianus and A. coeruleus live in shallower waters and are not as tolerant of silt as A. chirurgus.
Although many groups show their maximum diversity in the Indo-West Pacific, a few show maximum diversity in the Americas. Two-thirds of the species of toadfishes, Batrachoididae, occur in New World waters (Collette & Russo 1981). The most generalized subfamily, the Batrachoidinae, is worldwide. However, the two most specialized subfamilies, the luminous midshipmen (Porichthyinae) and the venomous toadfishes (Thalassophryninae), are restricted to the western Atlantic and eastern Pacific (plus a few freshwater species derived from Atlantic or Pacific marine species).
Eastern Pacific region
The eastern Pacific region contains another radiation related to and only recently separated from the western Atlantic. The region contains only four to eight genera of zooxanthellate corals (Rosen 1988) and fewer species of fishes than are present in the western Atlantic. Some widespread taxa, such as the Bluefish (Pomatomus saltatrix) and Cobia (Rachycentron canadum), are absent. The eastern Pacific barrier, the huge expanse of open water between the central and south Pacific islands and the American mainland, acts as a distance barrier limiting the movement of 86% of shore species from the central Pacific (Briggs 1974).
Elevation of the Panamanian Isthmus approximately 3 million years ago separated the continuous distribution of species into eastern Pacific and western Atlantic populations. David Starr Jordan, pioneer American ichthyologist, referred to such pairs of species as geminate species, related species divided by the isthmus, such as the Spanish mackerels, Scomberomorus sierra, in the eastern Pacific, and S. brasiliensis, in the Caribbean Sea. Some geminate species have clearly differentiated into what can be called good species from a morphological and sometimes from a genetic point of view as well, such as Spanish mackerels and toadfishes of the genus Batrachoides. Others, such as the halfbeaks of the genus Hyporhamphus are less well differentiated morphologically, making molecular methods useful to reach decisions on the status of the populations on either side of the isthmus (see The Panama barrier).
The completeness of the eastern Pacific barrier is, however, not quite as distinct as Briggs (1974) implied (Lessios & Robertson 2006). Several Indo-West Pacific shore fishes actually cross the eastern Pacific barrier and are found at offshore islands such as the Revilligedos off the coast of Mexico, and Clipperton and Cocos off the coast of Costa Rica. Distributions of a species of mackerel (Scomber australasicus) and a needlefish (Tylosurus acus melanotus) extend from the western Pacific through the Hawaiian Islands to these islands, but these species are replaced by related forms (S. japonicus and T. Pacificus, respectively) along the eastern Pacific coast of Middle America. These exceptions to the completeness of the eastern Pacific barrier may be related to habitat differences between the offshore islands and the mainland.
The Panama Canal connects the eastern Pacific with the western Atlantic. However, unlike the Suez Canal, the Panama Canal is not at sea level. It contains a freshwater lake, Lake Gatun, in its middle, and fresh water is used to raise the water level in a series of locks to lift ships up to the lake and then down to the ocean on the other side. This freshwater barrier prevents marine species from moving between the two oceans, with the exception of a few species that tolerate a wide range of salinities (McCosker & Dawson 1975). A proposed sea-level canal would allow mixing of the two different faunas and might have grave effects on the fishes and marine invertebrates on both sides of the isthmus. Diseases, parasites, and aggressive Indo-West Pacific species that pose little current danger in the eastern Pacific, such as the crown-of-thorns starfish and a sea snake, might do severe damage to coral reefs and the fish fauna of the western Atlantic (Briggs 1974).
Eastern Atlantic region
In the eastern Atlantic Ocean, tropical shore fishes are restricted to the Gulf of Guinea, a relatively small area that extends from Dakar, Senegal, to Angola, and includes offshore islands such as the Cape Verde Islands, Annobón, and Fernando Po. Coral cover is sparse in the tropical part of the eastern Atlantic, partly due to the large amount of freshwater runoff and accompanying sediment that flows out of such rivers as the Congo, Niger, and Volta. Only a few eastern Atlantic localities have as many as four to eight genera of zooxanthellate corals, other localities having only one to three genera (Rosen 1988). The eastern Atlantic is depauperate in many fish and invertebrate groups and contains only about 500 species of shore fishes. A few families, such as the porgies (Sparidae), have radiated in the eastern Atlantic.
Comparisons among genera of shore fishes of the western Atlantic, eastern Pacific, and eastern Atlantic demonstrate the relative depauperate nature of many eastern Atlantic groups. For example, four genera that contain two to four species each in the western Atlantic and eastern Pacific have only a single species in the eastern Atlantic (Table 16.1). Such comparison of patterns of diversity among different families can tell us much about not only the zoogeography of a group but also its probable phylogenetic history, particularly if we apply modern approaches to both zoogeography and phylogeny.
Table 16.1
Numbers of species in selected genera of inshore fishes from the western Atlantic, estern Pacific, and Gulf of Guinea (numbers in parentheses indicate freshwater species of marine origin).
Mediterranean Sea
The Mediterranean Sea is a somewhat depauperate part of the eastern Atlantic Ocean, with about 540 species of fishes. Drying out during the Messinian Salinity Crisis millions of years ago eliminated most fishes from the Mediterranean, and cooler temperatures in the Straits of Gibraltar prevented warm water fishes found in the Gulf of Guinea from moving into the warm waters of the eastern Mediterranean
In 1869, a sea-level route, the Suez Canal, was opened, connecting the warm but depauperate eastern Mediterranean with the Red Sea, the latter being part of the rich Indo-West Pacific region. For some time after construction, faunal transfers between the Red Sea and the Mediterranean were inhibited by the saline waters of the Bitter Lakes in the middle of the canal. In the first edition of A history of fishes in 1931, Norman reported that 16 species of Red Sea marine fishes had moved through the Suez Canal and established themselves in the eastern Mediterranean Sea. In the ensuing decades, Adam Ben-Tuvia, Daniel Golani, and others raised the number to 24, 27, 31, 46, and most recently to 68 (Golani 2006). All but one of the species (a seabass, Dicentrarchus) are what are termed Lessepsian migrants (named after Ferdinand Lesseps who was in charge of constructing the canal), having moved in one direction, from the Red Sea into the eastern Mediterranean. At least 24 species of Red Sea origin have reached as far west as the southeastern Aegean Sea. As an example of how successful a migrant can be, the Brushtooth Lizardfish (Saurida undosquamis) was first taken in the Mediterranean in 1952. By 1955, 266 tons of this lizardfish were landed by local trawlers, constituting close to 20% of the trawler catch in Israeli waters (Golani 1993).
Why have these movements been virtually onedirectional? First, the diversity of inshore fishes is greater in the Red Sea, part of the Indo-West Pacific fish fauna, than in the Mediterranean, suggesting that niches are more completely filled in the Red Sea, which means fewer ecological opportunities for new immigrants. Second, there appears to be an “empty niche” in the eastern Mediterranean, associated with water temperatures, with temperatures again being warm enough for warm water fishes.
Finally, many of the species that penetrated the canal are widespread species, adapted to a wide variety of living conditions. Consider the distributions of three of the invading species, a halfbeak and two mackerels. Hemiramphus far is the most widespread member of its genus, known from South Africa across the Indian Ocean north to Okinawa, south to Australia, and east to Tonga and Fiji. It, rather than the Red Sea-Persian Gulf He. marginatus, successfully moved through the canal and established populations that have now spread west and north as far as Albania. The two mackerels, the Narrow-barred Spanish Mackerel (Scomberomorus commerson) and the Indian Mackerel (Rastrelliger kanagurta), are the most widespread members of their genera, occurring from South Africa north to the Red Sea, east to China and Japan, and south to Australia and Fiji (Collette & Nauen 1983, maps on pp. 49 and 63). Such generally successful colonist species could have been predicted as the most likely taxa to take advantage of the opportunities in the eastern Mediterranean because they are adapted to a wide range of ecological conditions.
Arctic and Antarctic fishes
The diversity and adaptations of fishes of the far north and south are treated (Polar regions). Marine shore and continental shelf species down to 200 m from Arctic and Antarctic waters account for about 5.6% of the total fish fauna. The two polar regions contain 538 species of fishes, 289 in the Arctic and 252 in Antarctica (Møller et al. 2005). Only 12 of 214 polar fish genera and 10 of 72 polar fish families are found in both areas. The Arctic region north of 60° in the Pacific (approximately Nunivak Island, Alaska) to Newfoundland and northern Norway in the Atlantic has 20–25% endemism (Briggs 1974) and contains 416 species in 96 families (Eastman 1997). Six groups dominate, comprising 58% of the fish fauna: zoarcoids, gadiforms, cottids, salmonids, pleuronectiforms, and chondrichthyans. Other groups include skates, herrings, greenlings, poachers, snailfishes, pricklebacks, wolffishes, and gunnels. Most of these groups have higher species diversity in the Pacific than in the Atlantic portions of the region (Briggs 1974, 1995).
Antarctica and the surrounding Southern Ocean contain 322 species of fishes in 50 families (Eastman 2005). The immediate Antarctic region has 174 species in 13 families, 88% of which are endemic. Antarctica has a higher level of endemism of fishes and invertebrates than the Arctic Ocean, although the Arctic contains 1.5 times the fish species and twice as many families (Briggs 1974). Of the fishes in the immediate Antarctic region, six families in the suborder Notothenioidei account for 55% of the species and more than 90% of the individuals. Primitive notothenioids, such as the Bovichtidae, occur in southern hemisphere habitats of Australia, New Zealand, and South America. Some families occur in both Antarctica and the surrounding continents (Nototheniidae and Channichthyidae), some occur in Antarctica and nearby oceanic islands such as Las Malvinas (the Falklands) (Harpagiferidae), and one family (Bathydraconidae) is restricted to Antarctica. Of the notothenioids, 97% are Antarctic endemics; even 70% of the non-notothenioids are endemic. Six other families that contribute multiple species to the region are, in order of species diversity (Eastman 2005): snailfishes (70 species), eel pouts (24), skates (eight), and eel cods, deepsea cods, and southern flounders (four species each).
In some cool water species, such as the chub mackerels Scomber japonicus and S. colias, distributions are interrupted by low-latitude regions. Such species are considered to have antitropical distributions, in that they are present in temperate waters on either side of the equator (Hubbs 1952). Other cold water species show tropical submergence, that is, they continue their ranges into tropical regions by submerging, moving into deeper waters that are the same temperature as the cold waters of Arctic and Antarctic regions.
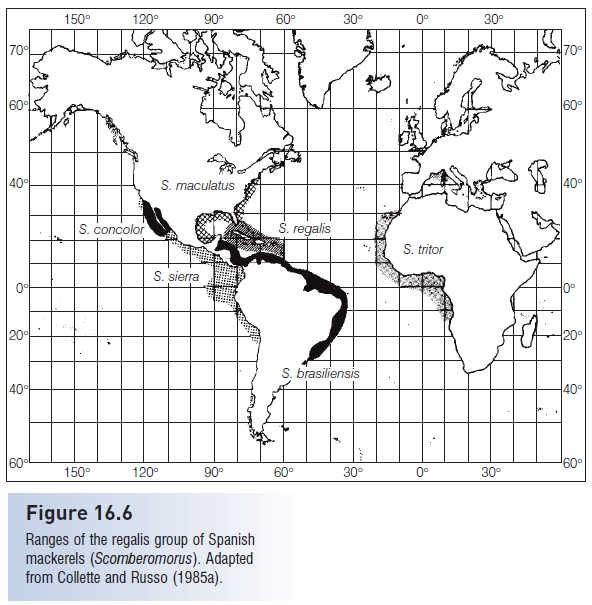
Figure 16.6
Ranges of the regalis group of Spanish mackerels (Scomberomorus). Adapted from Collette and Russo (1985a).
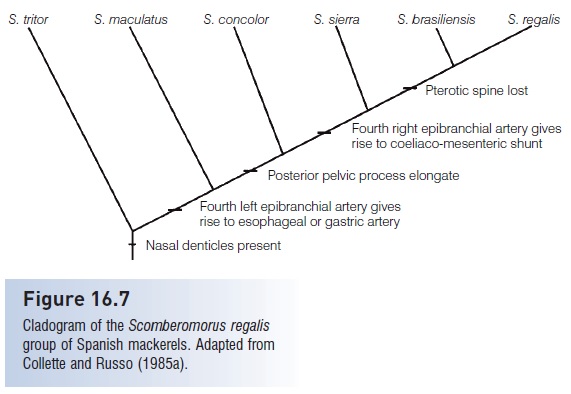
Figure 16.7
Cladogram of the Scomberomorus regalis group of Spanish mackerels. Adapted from Collette and Russo (1985a).
Related Topics