Chapter: Genetics and Molecular Biology: Generating Genetic Diversity: Antibodies
J Regions - Generating Genetic Diversity: Antibodies
The J Regions
The segments of embryonic DNA encoding the V
regions of both the lambda and kappa light chains of mouse IgG do not code for
about 13 amino acids normally considered to be the last part of the variable
region of the protein. The sequence of the embryonic C region DNA does not
solve the problem, for it contains just the amino acids that are constant from
antibody to antibody. The missing amino acids have been found to be encoded by
a segment of DNA lying from 1 to 3 kilobases (Kb) upstream from the 5’ end of
the C region gene. This region is called the J region because it is joined to
the V region in the process of DNA rearrangement that connects a V and C region
pair.
Of course, the most direct demonstration of the
existence of a J region was provided by sequencing. But Tonegawa had first
performed R-loop-ing to locate the J region approximately. Light chain
messenger from myelomas was hybridized to C region DNA cloned from embryos
(Fig. 20.6). The R-looped structures of the lambda chain located a short region
with the necessary coding properties about 1,000 to 3,000 nu-cleotides upstream
from the C region. By contrast, kappa light chain mRNAs extracted from a series
of myelomas indicated the existence of four different J regions ahead of the
kappa C segment. Thus the presence of these J regions adds another means for
generation of antibody
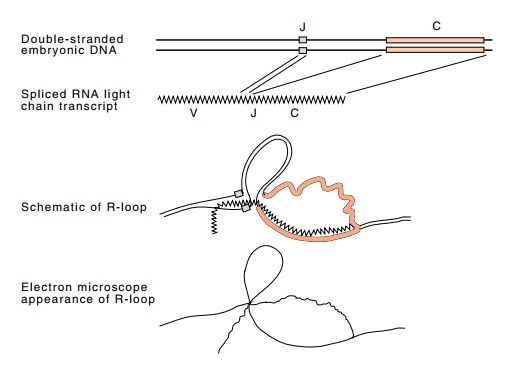
Figure
20.6 Genesis of an R-loop
demonstrating the existence and location ofa J region. In the embryonic DNA no
V region is near a J-C region.
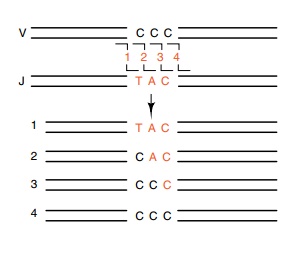
Figure
20.7 The variable positionof the
crossover joining the V and J regions can create several different codons at
the join region.
diversity. Any of the several hundred V segments
can be linked to the C segment by any one of the four J regions. Since the J
regions of the light chains lie just beyond the third hypervariable region of
the antibody, variability in this position has a direct effect on the structure
of the antigen-binding site of the resultant antibody and the affinity of the
antibody-antigen binding.
While nucleotide sequencing revealed a total of
five kappa chain J regions, sequencing of myeloma proteins showed that one of
the J regions is not functional. This finding is in keeping with the absence of
a GT sequence at the beginning of a splice site on this potential J region
gene.
Nucleotide sequencing by Leder demonstrated that additional diver-sity was also generated in connection with the J regions. The DNA crossover site that connects the V segment to one of the J regions is not fixed. Slight variability in its position creates codons not present on either the V or the J segment being joined (Fig. 20.7).
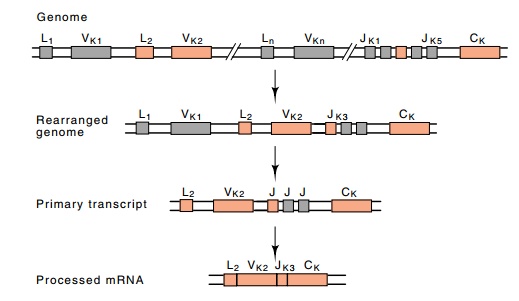
The J region and the surrounding sequences are
special. Immediately upstream of the J region itself lie signals that are used
in the DNA splicing of a V region to the J region (Fig. 20.8). Then,
immediately after the J region are the nucleotides that signal the mRNA
processing machinery so that the intervening sequence of about 2,500 bases
be-tween the J region and the beginning of the C region can be removed and the
messenger can be made continuous for translation. Later sections explain two
additional functions of this region: activating transcription and serving as
one end of a deletion that changes the C region joined to the V-J region.
Related Topics