Chapter: Psychology: The Genetic and Evolutionary Roots of Behavior
Genetics and DNA
GENETICS AND DNA
People often speak of the “genes
for obesity,” or the “genes for depression.” But what exactly is the linkage
between the complex molecules we call genes
and these large-scale observable traits? Let’s start by taking a look at what
genes are and how they operate.
Living things are all made of
cells, and the diversity of cells is
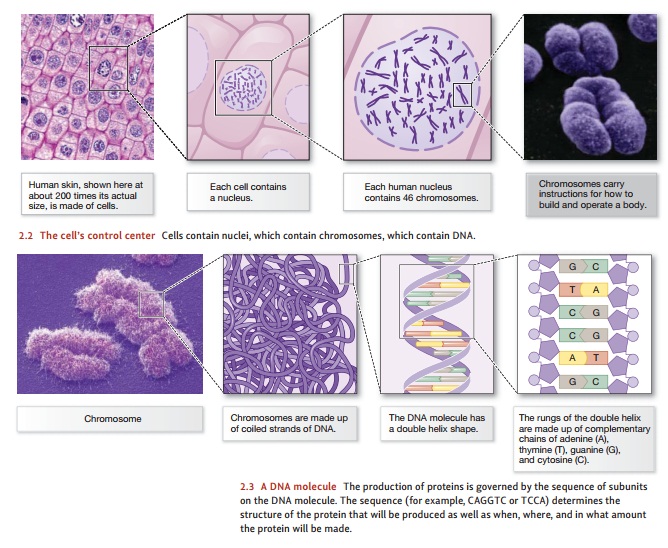
immense. For example, Figure 2.1 shows just three of the cell types found in the
human body—each cell type
with a different size, shape, and
function. What makes these cells so different is the pro-teins each type
contains, with each cell producing the specific proteins necessary for its role
in the body. But what governs the production of proteins? The answer lies in
the cell’s genes.
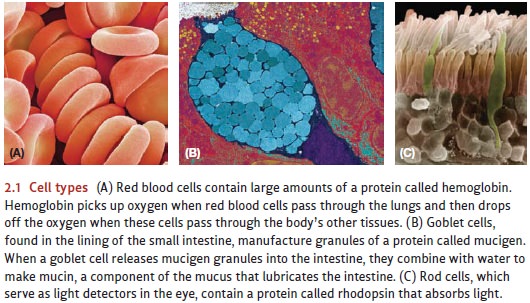
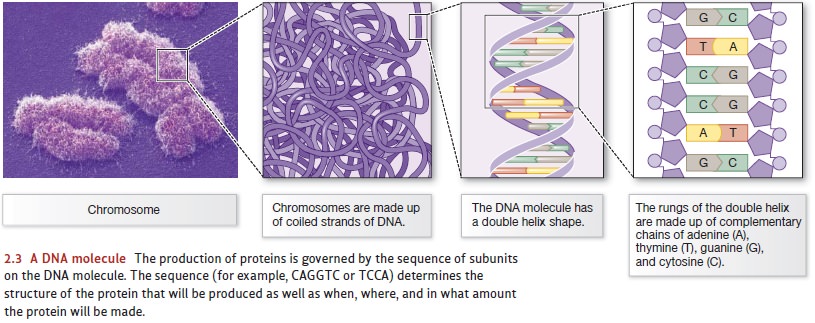
With just a few exceptions, all
cells in a plant or animal contain a nucleus—
essentially the biological control center for the cell (Figure 2.2). Within the
nucleus are the cell’s chromosomes,
complex structures that each contain a single long molecule of DNA (deoxyribonucleic acid). The DNA,
in turn, governs the cell’s structure and itschemical processes by providing a
set of detailed instructions for making the structural proteins that give each
cell its shape and for making the proteins called enzymes that govern the
cell’s functioning.
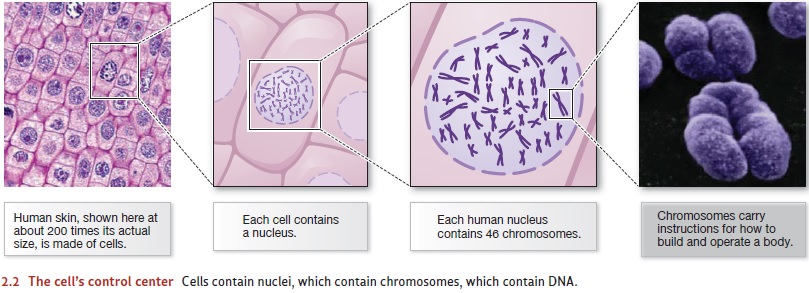
Genes
A DNA molecule is constructed like a twisted ladder, forming a shape known as a dou-ble helix. The rungs in the DNA ladder are made up of chemical subunits typically referred to by their single-letter abbreviations: A (adenine), T (thymine), C (cytosine), and G (guanine). Each rung consists of a pair of these subunits: A always pairs with T, and C always pairs with G.
The A–T and C–G pairs can be
placed in any sequence along a DNA molecule, just as beads of different colors
can be strung in any order along a necklace. This feature of DNA enables the
molecule to carry instructions for building proteins. If we read along one of
the uprights of the ladder, we encounter a sequence of letters (Figure 2.3;
from top to bottom along the right side of the molecule, the sequence is CAGGTC).
The machinery inside each cell “reads” this molecular sequence in sections;
some of the sec-tions describe the structure of a protein, while other sections
control when, where, and in what amount each protein will be made. The section
of a DNA molecule that describes the structure of a protein, together with its
control sequences, is called a gene.
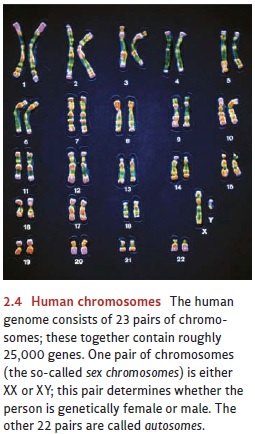
The DNA in each chromosome
encodes instructions for building hundreds of different proteins. In organisms
that reproduce sexually, these chromosomes come in pairs (Figure 2.4). Human
cells have 23 pairs, for a total of 46 chromosomes per cell; and each human
cell contains between 20,000 and 30,000 protein-coding genes (Stein, 2004;
Venter, Adams, Myers, Li, Mural et al., 2001). This count has astonished many
scientists, who had anticipated a number four or five times larger! Indeed, the
numbers surely seem small when we compare these counts to the 20,000 or so
genes in a simple roundworm (C. elegans)
cell. It’s important to bear in mind, though, that very few traits are
specified by single genes. Instead, what matters is the combination of genes,
and our (roughly) 25,000 genes allow for a lot of combinations.
Gene Expression
Virtually every cell in a person’s body contains a copy of the same 46 chromosomes, collectively called the person’s genome. Why then do the cells, all containing the same DNA, end up different from each other in their structure and functions (see Figure 2.1)? The answer lies in the fact that within each cell, the genes are not all active all the time. Instead, some genes in each cell are expressed and some are not. In fact, the process of gene expression is quite intricate, controlled by the interaction of an enormous variety of factors. In all cases, though, these factors work by modifying the biochemical environ-ment inside the cell, and it’s this intracellular environment that turns specific genes “on” or “off.”
One of the variables that can
alter the biochemistry within a cell is the environment just outside of the cell—and so whether a
gene will be expressed (and thus how the cell will grow) often depends on what
other cells are nearby. Another variable is timing— and so some genes are
active early in an organism’s development, but not later. Still other variables
depend on the organism’s overall environment and its behavior. For example, temperature influences gene expression,
and so does sheer stimulation—so that for some genes, expression in a complex,
engaging environment will be different from the expression in an impoverished
environment with little stimulation. Still other key factors are (perhaps
surprisingly) lodged in the social world—so that in some cases, gene expression
depends on the people you’re with and how you’re inter-acting with them (e.g.,
Cole, 2009). Other aspects of the organism’s behavior also matter: For example,
extensive exercise will cause a person to build larger muscles because exercise
triggers biochemical changes in the body that activate certain genes. These
genes then produce the proteins that become the building blocks of larger,
stronger muscles.
Notice, therefore, that how (or
whether) genes will be expressed depends on both the environment and
experience. As a result, it makes no sense to talk about “purely” genetic
effects. We need instead always to emphasize the interaction between genetic factors and the context, and we also
need to bear in mind that the interaction involves effects in both directions:
As we’ve just seen, the environmental setting and the organ-ism’s experiences
have a huge impact on gene expression; therefore, factors of experi-ence shape
how the genes operate. At the same time, genetic factors lead an organism to a
certain pattern of sensitivities and a certain capacity to remember. Therefore,
genetic factors shape what the organism experiences. As a result of all this,
the often-mentioned “nature/nurture” distinction, cleanly separating genetic
from environmen-tal effects, is misleading. Neither type of influence operates
without the other.
This interaction between
environment and genes also leads to another lesson: People often seem to assume
that an organism’s genes determine its destiny; a particu-lar genetic pattern,
they believe, will govern how the organism turns out. Instead, an organism’s
genes define only its genotype—the
specific sequence of genes on each of its chromosomes. In contrast, the
organism’s traits and capacities define its phenotype—what the organism is actually like. And the phenotype is
a product ofboththe genotype and the environmental context,
interacting in the ways we’ve discussed. Thus, the organism’s genotype provides
a crucial starting point, but does not by itself specify how the organism will
end up.
Gene Transmission
Why does an organism have the
genes that it does? The immediate answer lies in biolog-ical inheritance,
because organisms inherit their genotype from their parents. Bear in mind here
that chromosomes come in pairs, and, in most cell growth, all the pairs are
copied into the new cells. However, when women make egg cells and men make
sperm cells, each egg or sperm receives just one chromosome from each pair.
Thus, in humans, each egg and each sperm contains 23 chromosomes rather than
the normal complement of 46 (i.e., 23 pairs). When egg and sperm combine, the
resulting cell ends up with two sets of 23—one from the egg (and so from the
mother) and one from the sperm (and so from the father), and thus the correct
number of chromosomes (Figure 2.5).
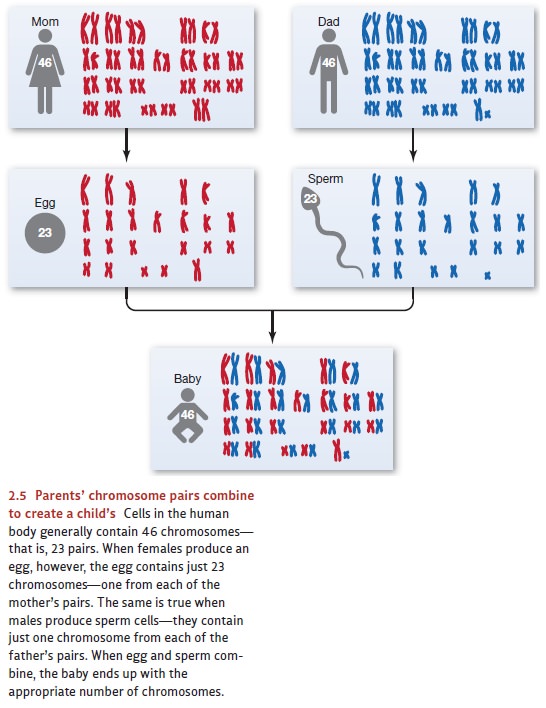
It’s a matter of chance, however,
which of the mother’s chromosomes (i.e., which chromosome from each of her
pairs) go into each egg, and this allows for a lot of possi-ble combinations
(more than 8 million). As a result, each egg is likely to be different from
every other. The same is true for the father—again it’s a matter of chance
which chromo-some from each of his pairs ends up in a particular sperm cell.
Then, it’s also largely chance that determines which of the father’s sperm
fertilizes which of the eggs. As a result of all of these chance occurrences,
each offspring is likely to represent a new com-bination of genes—a new play in
the genetic lottery we call sexual reproduction.
Interactions among Genes
Scientists are just beginning to
understand exactly how genes lead to a particular struc-ture—whether the
structure is a human heart with its four chambers, a human eye, or a human
brain ready to learn language. We do, however, know a lot about the broad
pat-terns of inheritance, and our modern understanding grows out of work done
more than a century ago: In the mid-1800s, a Moravian monk named Gregor Mendel
(Figure 2.6) was cultivating pea plants in a monastery garden in Brünn (now
Brno, Czech Republic). Mendel carefully observed the results of
cross-fertilizing one variety of plant with another, and from his observations
he worked out some basic laws of genetic inheritance. Mendel presented his ideas
in 1865 and 1866, first in a pair of lectures at the Brünn Natural History
Society and then in a paper published in the Brünn Society’s Proceedings.
However, his work was not widely appreciated, and Mendel’s extraordi-nary
achievement was not recognized until after 1900 (see Provine, 1971).

Let’s start, therefore, with the
same kind of case that Mendel started with: a character-istic shaped largely by
variation in just one gene. (We mentioned earlier that the huge majority of
genetic influences are more complex than this and depend on an intricate
interaction among many genes; still, a discussion of the single-gene case will
allow us to get some important fundamentals out in view.) We have said that
chromosomes come in pairs, and since genes
are located on
chromosomes, genes also come
in pairs: Each gene occupies a
specific position within its chromosome—called the gene’s locus—and for each gene,
there is a partner gene located at the corresponding locus on the other half of
the chromosomal pair. The two genes in each pair—one on the chromosome
contributed by the father, and one on the chromosome contributed by the
mother—may be identical, or they may differ. If the paired genes are identical,
we say that the individual is homozygous
for that gene; if the two are different, the
individual is heterozygous for
the gene. In
all cases, though, the variations
of a specific gene are referred to as alleles
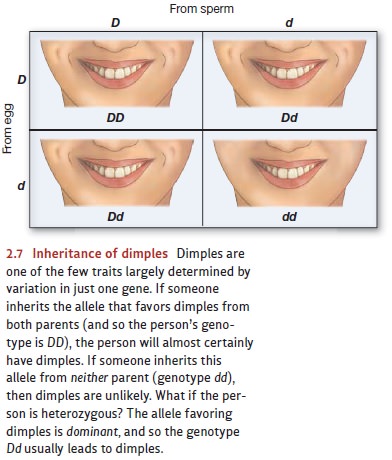
of that gene. How do things unfold if a person is heterozygous—having one
allele of a particular gene on one chromosome and a different allele of that
gene on the other chromosome? The simplest result is a relationship of dominance. As an illustration, consider dimples, one of
the few human
traits largely determined
by variation in
just one gene (Figure 2.7). If someone inherits from
both parents the allele that favors dimples, then the person is virtually
certain to end up with dimples. And if the person doesn’t inherit this allele
from either parent, then she won’t
have dimples. But what if the person inher- its the allele that favors dimples
from one parent, but not from the other? In this case, the person will probably
still have dimples because the allele for dimples is dominant: it will exert its effect whether the other member of the
gene pair is the same or not. In contrast, other alleles are recessive: A recessive allele will
affect the phenotype only if it matches the allele of its partner gene.
A handful of other human traits
are also based on a single gene pair; the list includes baldness, red-green
color blindness, and high susceptibility to poison ivy (all recessive).
Notice, though, that
these examples all
involve normal variations
among genes. Sometimes, however,
a person inherits a rare malfunctioning gene that can cause seri- ous medical
and psychological problems. One example is the gene (on chromosome 12) that
causes a disorder known as phenylketonuria
(PKU). This single (recessive) gene
can lead to profound mental retardation.
In fact, the linkage between PKU
and retardation is well understood. Because of the defective gene, someone with
PKU produces far too little of a specific digestive enzyme. As a result, the
person is unable to metabolize phenylalanine, a common amino acid. This leads
to a buildup
of undigested phenylalanine
in the body
and causes severe problems in brain development.
The treatment for PKU is
straightforward: If we ensure that there is little phenylalanine in the
person’s diet, then his inability to digest this chemical doesn’t matter at
all. In that case, the inherited disorder is inconsequential and the person
develops normally. Notice, then, that the PKU genotype may or may not lead
to the phenotype of
retardation; it all depends on the person’s diet. The alleles of a gene can also interact in other ways. In
some cases, the alleles are codominant—a relationship
in which both
genes in the
pair affect the
phenotype. A common example is blood type: If the person inherits the
allele favoring blood type A from one
parent and the
allele favoring blood
type B from
the other parent,
the person will end up with type AB—expressing both genes.
In other cases, the alleles are
in a relationship of incomplete dominance,
so that a per- son with two
different alleles will
have a phenotype
that’s intermediate between
the types favored by each allele on its own. An important example concerns
the serotonin transporter gene, located
on chromosome 17. Serotonin is one of the chemicals that nervecells use to
communicate with each other, and it plays a role (among other functions) in
regulating our emotions. Nerve cells that use serotonin also make a protein,
called the serotonin transporter, whose job it is to mop up serotonin after it
has been used to send a message. The gene that controls the production of this
protein has two alleles that differ in the length of one part of the gene. An
individual can have the longer allele on both of their copies of chromosome 17
(and so their genotype is described as l/l),
or the short allele on both (a genotype of s/s),
or the short allele on one and the long allele on the other (s/l).
The long allele of this gene is incompletely dominant over the short
allele. As a result, people with the l/l
genotype produce more serotonin transporter than do people with the s/l genotype; they, in turn, produce
more serotonin transporter than do people with the s/s genotype. How does this matter for the individual? Studies have
indicated that people who produce less of the serotonin transporter are at
greater risk for both depression and anxiety and may react differently to
antidepressant medication. Because these claims have been controversial (Arias,
Catalán, Gastó, Gutiérrez, & Fañanás, 2003; G. Brown & Harris, 2008;
Caspi, Sugden, Moffitt, Taylor, Craig et al., 2003; Lesch, Bengel, Heils,
Sabol, Greenberg et al., 1996; Munafo, Durrant, Lewis, & Flint, 2009;
Risch, Herrell, Lehner, Liang, Eaves et al., 2009), it’s plain that more
stud-ies are needed. Even so, there’s no question that serotonin plays a key
role in regulating our moods, appetites, and sleep schedules—facts that
guarantee continued research on the serotonin transporter gene.
Polygenic Inheritance
We have discussed several
examples of variations in a single gene, and we’ve also looked at some of the
ways that the alleles of a gene can interact. But keep in mind that the vast
majority of an organism’s traits are influenced by many genes, through a pattern known as polygenic inheritance. Examples here include bipolar disorder and
schizophrenia. Both disorders are much more likely if a person inherits a
certain genetic pattern, but the pattern includes many genes, each playing a
part in creating the disease risk.
The logic of polygenic
inheritance is, however, the same as the logic we’ve been dis-cussing: The
genes that are part of the polygenic pattern come in pairs, and the genes
within each pair may have the same alleles or different ones. If the genes are
different, then one allele may be dominant or (more likely) incompletely
dominant or codomi-nant. And, above all, the way that the genotype is reflected
in the person’s phenotype will depend on an interaction between the genetic
pattern and numerous factors in the person’s environment and experience.
Related Topics