Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Soft anatomy
Gas bladder of Fishes
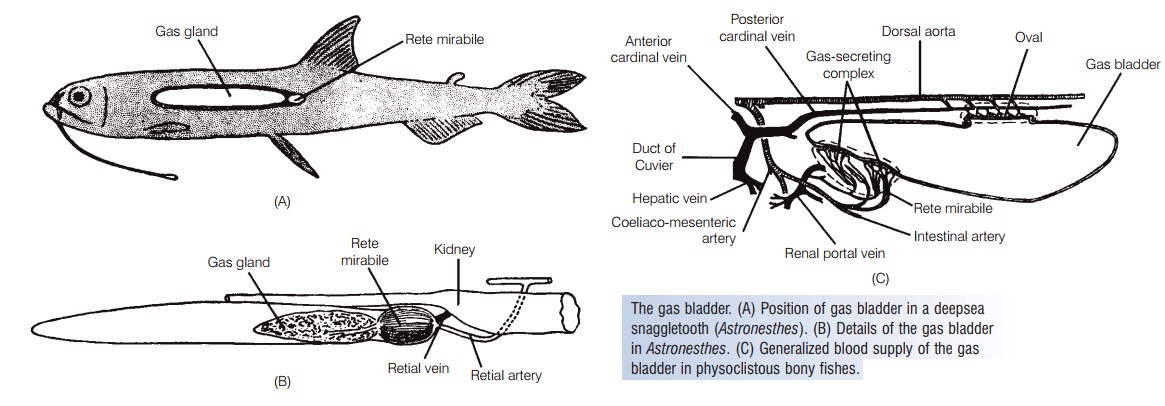
Gas bladder
The gas bladder (swim bladder) is a gas-filled sac located between the alimentary canal and the kidneys (Jones 1957;Marshall 1960). It is filled with carbon dioxide, oxygen, and nitrogen in different proportions than occur in air, making the term “air bladder” inappropriate. The original function of the gas bladder was probably as a lung, but inmost fishes today it functions mainly as a hydrostatic organ that helps control buoyancy. It also plays a role in respiration, sound production, and sound reception in some fishes. Some species in at least 79 of 425 families of extant teleost `shave lost their gas bladders, at least as adults (McCune &Carlson 2004). Most of these fishes are either benthic ordeep-sea species. Billfishes (Istiophoridae) and two genera of halfbeaks (all 10 species of Hemiramphus and one of two species of Oxyporhamphus) have a vesicular gas bladder composed of many discrete gas-filled vesicles (Tibbett set al. 2007).
Embryo logically, the gas bladder is a two-layered (tunicaexterna and tunica interna), specialized outgrowth of theroof of the foregut and possesses tissues similar to those of the foregut, as shown here:

The structures and mechanisms by which gases enter and are released from the gas bladder differ in the major groups ofteleost`s. Thepneumatic duct is a connection between the gas bladder and the gut. Physostomous fishes retainthe connection in adults, whereasphysoclistous fishes lose the connection in adults, if it is present at all during development. In physostomous fishes, gas can be taken inland emitted through the pneumatic duct. More primitive soft-rayed teleost`s have the primitive physostomous condition; whereas more advanced spiny-rayed fishes are physoclistous, lacking a pneumatic duct (see Buoyancy regulation).
Another, more complex mechanism, which involves two distinct regions of the gas bladder, has evolved to allow gas exchange in these fishes (Fig. 4.9). The anteroventral secretory regioncontains the gas gland and the rete mirabile.
The gas gland secretes lactic acid into the beginning of the capillary loop. This acidifies and reduces the solubility of all dissolved gases. A change of 1 pH unit releases 50% of the oxygen bound to hemoglobin. This raises the partial pressure of blood oxygen by the Root and Bohr effects (see, Gas transport).
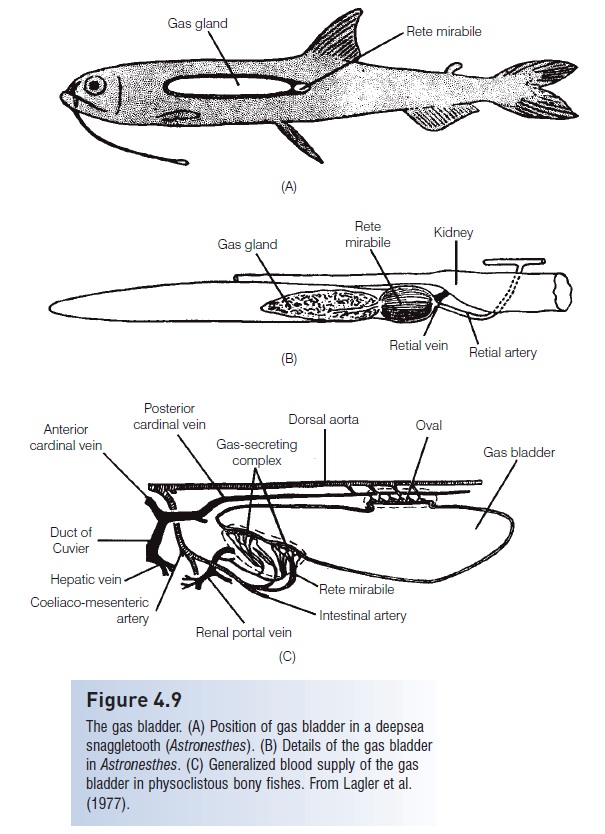
Figure 4.9
The gas bladder. (A) Position of gas bladder in a deepseasnaggletooth (Astronesthes). (B) Details of the gas bladder inAstronesthes. (C) Generalized blood supply of the gas bladder in physoclistous bony fishes. From Lagler et al.(1977).
The rete mirabile, or wonder net, is not actually a net but a looping bundle of arterial and venous capillaries associated with the gas gland that functions as a countercurrent multiplier. The rete is better developed in deep dwellingfishes that have longer retial capillaries, thus providing more surface area and allowing a greater multiplying factor. Rattails (Macrouridae) and ophidioids living at abyssal depths of 4000 m and deeper have retial capillaries25 mm in length or more; shallow water forms haveretes only 1 mm long (Marshall 1971).
The posterodorsal resorptive region of the gas bladder is called the oval. It develops from the distal end of thedegenerating pneumatic duct and consists of a thin, highly vascularized area. Circular muscles contract and close off the oval, preventing outflow of gases. Longitudinal muscles contract and expose the oval, permitting gas escape. The walls of the gas bladder are lined with a layer of cells containing crystals of guanine 3 μm thick, which decreases permeability by 40 times over an unlined membrane and thus limits gas escape except at the oval, when it is open.
The gas bladder of physostomous fishes receives blood from a branch of the coeliaco-mesenteric artery. Blood isreturned to the heart through the hepatic portal system. The rete, oval, and gas bladder wall of physicists are supplied by the coeliaco-mesenteric artery and blood is returned by a vein from the hepatic portal system. The oval and bladder wall are also supplied by intercostal branches of the dorsal aorta and returned through the post cardinal system.
Nervous innervation of the gas bladder is sympathetic through a branch from the coeliaco-mesenteric ganglion and by branches of the left and right intestinal vagus (X)nerves. Cutting the vagus prevents gas secretion into thegas bladder. Gas secretion is also inhibited by atropine, a
cholinesterase blocker. The gas gland has high cholinesteraseactivity, and the secretory fibers are probably cholinergic. Sensory nerve endings function as stretch receptors,responding to stretching or slackening of the gas bladder,thus providing information to the fish about the relativefullness of the gas bladder.
Related Topics