Chapter: Plant Biology : Seed plants
Evolution of flowering plants
EVOLUTION OF FLOWERING PLANTS
Key Notes
Origin of the flower
The fertile parts of a flower are a further reduction from those of other seed plants. The anther is simple and the male gametophyte consists of a pollen tube nucleus and two sperm nuclei. The ovule is enclosed by a carpel and the female gametophyte is reduced to an embryo sac. Fertilization of the endosperm is unique to angiosperms.
The earliest flowering plants
The earliest known fossils come from the early Cretaceous and resemble pollen grains of living Chloranthaceae. DNA evidence suggests that a shrub,Amborella, and water-lilies diverged first in angiosperm evolution. These plants share features and it is likely that the primitive flowers were mainly small but variable in size with few but indefinite numbers of fertile parts, some hermaphrodite, other unisexual. Some had a bract. A range of large and small flowers appeared soon after.
Early evolution
The sepals derive from a whorl of leaves around the fertile parts. Some petals derived from sterile stamens, others from a second whorl of sepals. There were two main lines of evolution, one to large flowers and specialist insect pollination, mainly by beetles, the other to small flowers in inflorescences pollinated by wind. Subsequent developments led to smaller flowers with floral parts in fives or threes.
Later evolution
In the late Cretaceous/early Tertiary period, flowering plants radiated to dominate the world, in conjunction with specialist flower-feeding insects. Innovations included fused petals, carpels inserted below the petals and bilateral symmetry. Some had inflorescences. Wind-pollination evolved many times. Most modern plant families appeared.
Specialization in flowers
A few plants are specialized to one or a few pollinator species. The orchids may attract just one species of bee and figs can only be visited by one group of wasps with which they have a close interdependent relationship. Generalization in pollination is much more common.
Evolution of vegetative structure
Flowering plants are enormously variable in vegetative structure, but the earliest were probably shrubs from which developed trees and herbaceous plants. Vessels appeared early but a few families have none. Dicot leaves are mainly net-veined, many monocot leaves parallel veined, but there are exceptions and some variable families.
Evolution of fruits and seeds
Primitive fruits are likely to have been fleshy and seeds filled with endosperm. Many fruit and seed types have evolved in parallel in different families. Major trends have been towards reduction in size of fruit and seed number, and towards seeds with a large embryo and little or no endosperm at maturity.
Classification
The distinctions between plant families are based mainly on flower form but there are often associated fruit and vegetative characters. Some large families vary little; some, particularly those with primitive features, vary more. The two main divisions, monocots and dicots, are confirmed by molecular evidence, except for those families that branched off early in evolution, now known as primitive dicots.
Three families of plants
As examples of features on which the classification is based, the daisies, grasses and orchids are all large important families each with many unifying features.
Origin of the flower
The flower of an angiosperm represents a further reduction and specialization of the heterosporous form discussed in relation to the other seed plants. The anther is simpler than that of the Bennettitales or Gnetales (Topics R1 and R3); the ‘fertile leaf’ is, in most, a simple stalk or filament. The male gametophyte is always reduced to a total of three cells . The origin of the carpel surrounding the ovules is obscure but it may be a modified leaf, or a cupule as in the Caytoniales . In nearly all angiosperms the ovary completely encloses the ovules. The carpel also has a stigma and usually a style joining this to the ovary. The carpel may have evolved in response to insect visitationof the flowers, perhaps as protection for the vulnerable and nutritious ovule. Insects could transfer pollen to other flowers and insect pollination is thought to be the primitive pollination type in angiosperms.
The female gametophyte that develops within the ovule, known as the embryo sac, is much reduced compared with other seed plants . No archegonia are produced and, in many, nuclei are formed without cell walls.
From pollination to fertilization in angiosperms takes a few hours and, in contrast to all other seed plants, the double fertilization leads to growth of the endosperm as the seed’s food store. This means that the food store only grows in fertilized seeds, potentially saving resources. In all other seed plants the female gametophyte provides the food store.
The earliest flowering plants
The earliest evidence of flowering plants is fossil pollen grains and fragmentary flowers and fruits from lower Cretaceous rocks, about 110 million years old (some 130 million year old fragments may be angiosperms), suggesting that flowering plants evolved some time in the Jurassic, perhaps 150 million years ago, long after the other seed plant groups . The fossil record is always fragmentary and the conditions for preservation such that only certain plants are likely to be preserved, e.g. those living by water or acid peat.
Small or isolated plant populations, which are likely to be particularly significant in evolution are not likely to be well preserved.
The pollen and form of the flower of the earliest fossils are similar to those of living members of the Chloranthaceae, a small family from tropical America, east Asia and New Zealand. The flowers of these plants are simple and tiny, up to 3 mm long, some genera bearing hermaphrodite flowers, others unisexual, borne in inflorescences (Fig. 1). Each flower contains one or three stamens and/or one carpel and some have a bract (small modified leaf) under them. They have no petals or sepals. Hermaphrodite species are pollinated by unspecialized insects; unisexual species by the wind. Several other living plant families are represented as fossils in slightly younger rocks, with a wide range of flower structure from large-flowered magnolias to catkin-bearing trees such as the planes. In some of these the flower structure is not rigidly set, and they have variable numbers of stamens and carpels surrounded by a perianth, i.e. modified leaves or bracts not clearly sepals or petals.
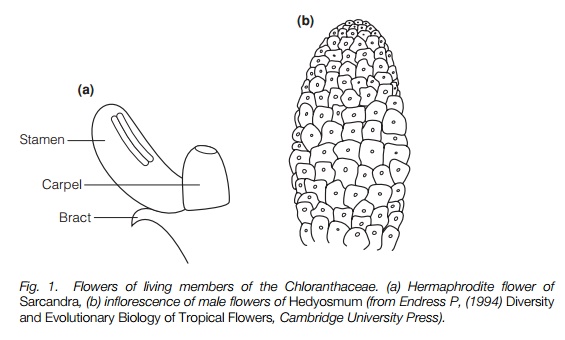
Over the last few years, DNA from genes in the nucleus, plasmids and mitochondria of many angiosperms has been analyzed. Putting all the molecular evidence together it appears that a shrub, Amborella, from New Caledonia that bears unisexual flowers, and the water-lilies, which are herbaceous aquatics bearing large hermaphrodite flowers, diverged from the other angiosperms earlier than any other group. Both of these have numerous stamens and an indefinite number of perianth segments. The Chloranthaceae and its relatives are, on this evidence, closer to other families.
Whatever the most primitive plant is, it is likely that the earliest flowers had certain features since these are shared by all the plants mentioned above and are consistent with the fossil record. There was a variable number of stamens and carpels, each separate and inserted above the perianth where this was present. Unisexual and hermaphrodite forms appeared early, and some had a perianth or bracts underneath the fertile parts but no well-defined petals or sepals. They are likely to have been visited and pollinated by unspecialized beetles, flies and wasps, though wind-pollination probably arose early on. They were small shrubs and perhaps herbs among dominant gymnosperms.
Early evolution
Adaptive radiation of flowering plants, i.e. their spread and diversification, occurred through the Cretaceous period in parallel with other seed plant groups and some living families appeared. Outer whorls of the flower became differentiated early on, the outermost layer, referred to as sepals even if there is onlyone whorl, probably evolving from bracts. When there are two different whorls, the inner whorl is referred to as the petals, but some probably evolved from a second whorl of sepals, others from stamens losing their anthers and becoming organs purely for attracting insects. Some water lilies have intermediate organs, stamens with flattened colored filaments, between the fertile stamens and larger entirely sterile petals. Cultivated ‘double’ flowers such as roses have some stamens replaced by petals . One main early line of evolution was towards a fixed number in each whorl, often five in the dicots and three in the monocots, making the flowers radially symmetrical (actinomorphic).
There was specialization for pollination, by beetles along one line of evolution including the magnolias, and by wind along another leading to the catkin-bearing trees. The insect-pollinated group had hermaphrodite flowers of, at first, increasing size and complexity, though later insect-pollinated flowers are smaller. By the end of the Cretaceous period, many families with actinomorphic insectpollinated flowers had appeared, such as buttercups, pinks and heathers. The wind-pollinated group retained small unisexual flowers, with few parts borne in inflorescences, usually with male and female on the same plant. This monoeciousbreeding system includes less than 5% of living species but several of these are dominants such as the oaks and beeches, birches, hazels and planes.
Later evolution
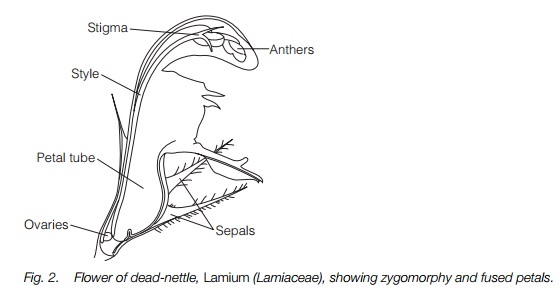
At the end of the Cretaceous and beginning of the Tertiary period, between 75 and 50 million years ago, there was a second and much larger adaptive radiation which saw the increasing dominance of the angiosperms and the appearance of the majority of modern plant families. This was closely associated with the adaptive radiation of specialist insect pollinators and leaf feeders, the butterflies and moths, long-tongued flies and bees. Innovations included the fusion of the lower part of the petals and/or the sepals into a tube and the fusion of the carpels. Bilaterally symmetrical, orzygomorphic, flowers appeared, usually facing sideways rather than upwards and with the lower petal(s) forming a lip as a landing platform for insects. Some flowers such as members of the thyme family have fused petals and zygomorphic flowers (Fig. 2). These were adaptations for specialist insect pollination and at least in part serve to exclude other visitors. Adaptations for pollination by birds or bats appeared later, mainly derived from specialist insect-pollinated flowers. The main trends are outlined in Table 1.

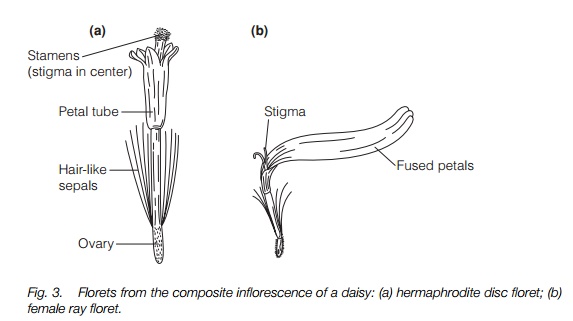
Aggregation of flowers into inflorescences is an adaptive trend in many plant families. It is taken to extreme specialization in the daisy family in which there are often two different forms of flower in each inflorescence, the whole aggregation, sometimes of more than 100 flowers, resembling a single flower (Fig. 3).
Specialization for wind-pollination has nearly always involved a reduction in size and numbers of floral parts. Some families are nearly all wind-pollinated, e.g. grasses, but wind-pollinated species occur in numerous mainly insectpollinated families and have evolved many times. Evolution in the reverse direction, from wind to insect-pollination has rarely occurred.
Specialization in flowers
A few flowers show extremes of specialization in their pollination. In manyorchids, each flower needs a single visit for the successful pollination of hundreds of seeds, so they can become highly selective in the species that they use. Some tropical species have a long spur growing out of the back of the flower with nectar at its base, which only moths with extremely long thin tongues can reach. The European bee orchid group, and a few other groups of orchids in other parts of the world, have flowers that resemble the female bee, both in looks and in scent, each orchid species resembling a different bee; they are visited and successfully pollinated by the males of that bee species.
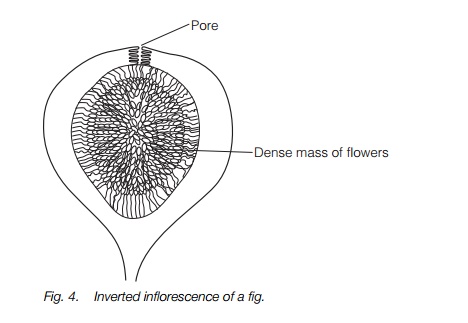
The figs, a large and important group of tropical trees and ‘stranglers’ have flowers in an inverted inflorescence with flowers opening into a central cavity, surrounded by a solid mass (Fig. 4). The inflorescences can only be penetrated by small wasps through a hole at the tip. The wasps lay their eggs, grow and mate in the inflorescences before collecting pollen and dispersing to another fig. Plant and wasp are totally interdependent. There is a similar interdependence between yuccas and moths, but otherwise such specialization is rare.
More generalist flowers are much more common than the specialists, particularly among the dominant vegetation, which are usually pollinated either by wind or by a range of insects and sometimes birds as well. Pollinating insects (and other animals) can vary manyfold in numbers from year to year and place to place, so specialization to one or a few species can be risky.
Evolution of vegetative structure
There is a great range of vegetative form from tall long-lived trees (although none grows as tall or lives as long as certain conifers) to short-lived plants . They may float in fresh water without attachment to any soil or spread by underground rhizomes to cover a large area. Their leaves range from minute duckweeds measuring around 1 mm in diameter, to some palm leaves that can exceed 15 m in length, or they may be reduced to spines or absent. Most woody plants are dicots and have secondary thickening . Palms, bamboos and some other monocots have numerous separate vascular bundles and most only start to produce a trunk when the bud has grown broad at ground level; their trunk often tapers only slightly or not at all. In a few monocot trees, the vascular bundles line up and they have some secondary thickening. Evidence for the direction of evolution in these plants is thin, and vegetative form is flexible, many families containing a wide variety of form and leaf shape. The earliest flowering plants and their living representatives are mainly shrubs or trees and all living gymnosperms are woody. It is likely that herbaceous plants are derived from woody forms originally, but some woody plants, including monocot trees, may be secondarily evolved from herbaceous plants. Short-lived plants are the end-points of the reduction.
Xylem vessels evolved early on, but a few angiosperms including Amborella and the Winteraceae, a family with flowers retaining many primitive features,have none and in water-lilies the xylem cells are somewhat intermediate between tracheids and vessels.
Dicot leaves are mostly net-veined, the veins joining together in many places. Among monocots there is great variation, although many have spear-shaped leaves with parallel veins. Some monocot families, especially the arum family, have enormous variation in leaf shape, with net venation like the dicots and some even developing holes in the leaf such as the ‘swiss-cheese plant’, Monstera, popular as a house plant.
Evolution of fruits and seeds
There is some dispute over which fruit type is primitive; fossil evidence gives few clues, but it is likely that the early fruits were fleshy as many of the representatives of primitive groups have fleshy fruits, sometimes with outgrowths
from the seed (an aril). Trends in the evolution of fruits have gone in many directions, towards larger or smaller fruits or seeds, and towards dry dehiscent fruits, but one main trend appears to have been for a reduction in
size and in number of seeds per fruit. There has been a parallel evolution of fruit types in many plant families, i.e. evolution along similar paths in unrelated groups.
Primitive seeds are likely to have been fairly large and rich in endosperm, features retained in some specialized large-seeded plants such as the coconuts. The small seeds, mainly filled by the embryo, that are seen in many families are likely to be derived. Some of the most successful families such as the grass and daisy families have one-seeded fruits, but another large family, the orchids, have many hundreds or thousands of tiny seeds per pod.
Classification
Classification of flowering plants is mainly based on the form of the flower, as this is regarded as a conservative character (one that does not change fast in evolution; often a suite of connected characters). Frequently it is correlated with the structure of fruits and vegetative form. Many plant families are defined by the shape and orientation of the flowers, and some large families have remarkably uniform flowers and growth form. Some families, particularly those with primitive features, are more varied but classified together because of a suite of characters, e.g. the buttercup family. Some of the relationships between families are obscure, particularly among those with primitive features.
Flowering plants have long been classified as two main divisions based on morphological characters: dicots, comprising about 70% of angiosperms, and monocots comprising the other 30%. The groupings are largely supported by molecular studies into their relationships, except for the group of families that retain primitive features, comprising about 5% of angiosperm speciesincluding water-lilies, magnolias and Chloranthaceae. These form a disparate group of more or less related families that diverged from the main angiosperm line earlier than the monocot/dicot split. They are now known as primitive dicots
Three families of plants
To illustrate the unifying features that are used to place plants in different families, examples of three large and important families are given.
Daisy family (Asteraceae or Compositae)
The daisy family has over 20 000 species, all with specialized inflorescences of tiny flowers (Fig. 3) each maturing one seed. They are unspecialized in their pollination but many are extremely attractive to bees, butterflies and other flower visitors. Fruits are dry and small and many are effectively dispersed by the wind using a cottony sail. They are mostly herbaceous with very few small trees, perhaps primitive in the family. They are associated particularly with dry regions and many have a deep tap root, but they flourish everywhere.
Grass family (Poaceae or Gramineae)
The grass family has about 9000 species and dominates large areas of the world. Grasses are entirely wind-pollinated except for one or two secondarily adapted to insect pollination. The fruit is one-seeded and dry, dispersed by wind or sticking to animals. All are herbaceous except for the bamboo group which havea most unusual woody form. Growth is not from the stem tip but from nodes on the stem allowing particular tolerance of grazing and fire .
Orchid family (Orchidaceae)
The orchid family has over 25 000 species. Their flowers have three sepals and three petals, one looking different from the others forming a lip. Their pollen is dispersed in aggregations (pollinia), and many species are adapted to one or a few insect species, usually bees. Many provide unusual food rewards such as oil or scent, or no food reward, deceiving their pollinators. The fruit is a pod with huge numbers of microscopic seeds deriving from one or few pollinations. All are herbaceous with specialist mycorrhizal infection in their roots and many are epiphytic .
Related Topics