Chapter: Anatomy of Flowering Plants: An Introduction to Structure and Development : Flower
Embryo Sac
Embryo Sac
Within
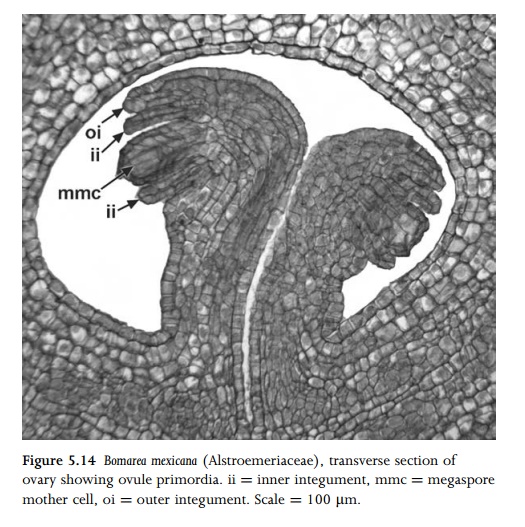
the nucellus, a single (normally hypodermal) cell becomes a primary sporogenous
cell (archesporial cell, or archespore). The archespore rarely consists of more
than one cell, though it can be multicellular in a few species (e.g. Brassica
campestris86), in which one cell produces the megagametophyte. In
turn, the archesporial cell either gives rise directly to the megaspore mother
cell (megasporocyte) or undergoes mitosis to form a primary parietal cell and a
megasporocyte. The megasporocyte then under-goes two meiotic divisions
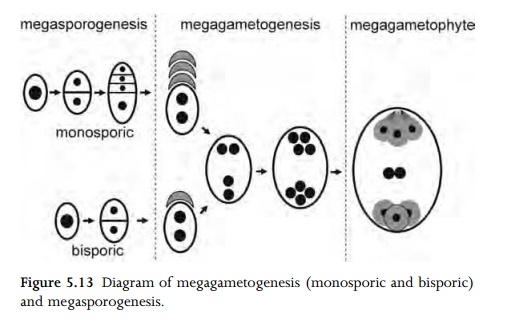
(megasporogenesis) to form a tetrad of four megaspores, which are usually
either in a linear or T-shaped arrangement. In the majority of angiosperm
flowers, one megaspore (most commonly the chalazal one) gives rise to the
mature embryo sac by further mitotic divisions, and the other three megaspores
degenerate (Fig. 5.13). This type of development is termed monosporic. However,
in relatively few angiosperms, two or four megaspores play a role in embryosac
formation; these types are termed bisporic or tetrasporic respectively69,125,126.
Degenerated megaspores are often sur-rounded by persistent callose.
In most
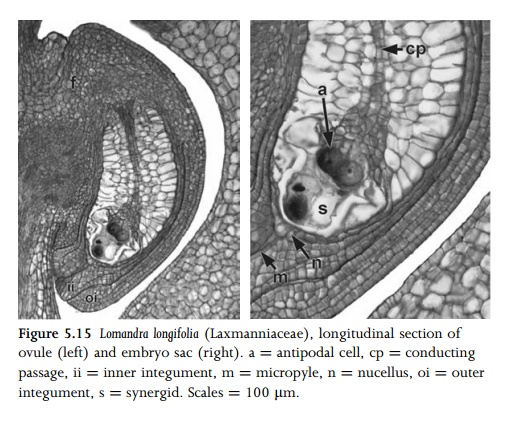
angiosperms the mature embryo sac (megagameto-phyte) possesses eight nuclei
arranged in seven cells (Fig. 5.15), though types with four and sixteen or more
nuclei have also been recorded. The most common type is monosporic and
eight-nucleate; this is sometimes termed the Polygonum type of embryo sac
development. At the binucleate stage, the two nuclei migrate to the micropylar
and chalazal poles and subsequently divide. Of the two micropylar nuclei, the
one closest to the micropyle divides to form the synergids, and the other
divides to form the egg cell and one of the polar nuclei. The two chalazal
nuclei each divide so that one forms two antipodal cells and the other forms an
antipodal and a polar nucleus. The two polar nuclei migrate to the centre and
fuse to form a diploid fusion nucleus. Cellularization follows, so that the
mature megagametophyte consists of three antipodal cells at the chalazal end, a
central cell with a fusion nucleus, and two synergids plus an egg cell at the
micropylar end.
The
synergids and the egg cell are so tightly pressed together that they are
collectively termed the egg apparatus. The syner-gids play a role in directing
the pollen tube into the embryo sac; they are calcium-rich and normally possess
a series of wall thickenings, the filiform apparatus, which extends into the
micro-pyle (Fig. 5.15). In many species the antipodals degenerate at an early
stage, but in others they persist, and sometimes undergo cell division (e.g. in
many grasses) or endoreduplication.
Related Topics