Chapter: Genetics and Molecular Biology: Attenuation and the trp Operon
Coupling Translation to Termination
Coupling Translation to Termination
The location of the leader peptide with respect to
the four different regions capable of hybrid formation in the trp leader region provides a simple
mechanism for regulating termination. If the 3-4 hybrid forms during
transcription of this region, termination is possible because this hybrid is
the “termination” loop. Conversely, if the 2-3 hybrid forms during
transcription of leader, then formation of the “termination” loop is prevented.
Finally, if the 1-2 hybrid forms, then region 2 is not available for formation
of the 2-3 hybrid, but as RNA polymerase transcribes regions 3 and 4, they are free
to base pair, and transcription terminates.
How can the presence or absence of tryptophan
affect formation of the 1-2 or 2-3 hybrid? Ribosomes translating the leader
region in the absence of charged tRNAs will stall at the trp codons and, owing to their location, they will block the
formation of the 1-2 hybrid (Fig. 13.12). Formation of the 2-3 hybrid is not
blocked, and as soon as these regions of the mRNA are synthesized, this hybrid
forms. Consequently, the 3-4 hybrid does not form in time to terminate
transcription, and termina-tion does not occur.
In the absence of any protein synthesis, ribosomes
cannot bind to the leader region, and the 1-2 hybrid forms. In turn, this
permits formation of the 3-4 hybrid. Termination follows. This happens in an in vitro
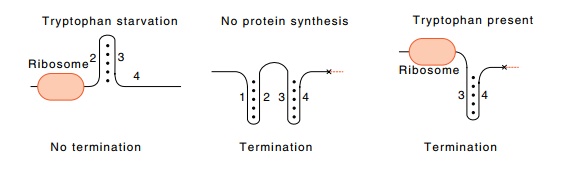
Figure
13.12 Possible structures of the leader
mRNA in the presence of ribo-somes under the three conditions: tryptophan
starvation, no protein synthesis, and tryptophan excess.

Finally, what happens in the presence of excess
tryptophan? In cells containing adequate levels of tryptophan, much initiation,
of course, will be blocked by the trp
repressor. The transcription that is initiated however, will be largely
terminated, for ribosomes either complete translation of the leader peptide and
permit hybrids 1-2 and 3-4 to form, or ribosomes remain awhile at the
termination codon. From this posi - tion they block formation of the 1-2
hybrid. In either case, the 3-4 hybrids form, and termination at the attenuator
occurs.
It is necessary to note that the relative
thermodynamic stability of the various leader hybrids is not important to
attenuation. The factor determining whether termination will occur is which
structures are not blocked from forming. The kinetics of formation of the
base-paired structures should be on time scales less than milliseconds so that
if they are not blocked from forming, they should form while the RNA is being
synthesized. Then several seconds after initiating transcription, RNA
polymerase reaches the attenuation site and terminates or not depend-ing on
whether or not the 3-4 hybrid has formed. The interval between initiation and
termination is much shorter than the interconversion time of many hybrid
structures, and they therefore will not necessarily have had time to adopt
their lowest-energy conformation (Table 13.1). The factor determining
termination is which of the hybrid structures exists at the time RNA polymerase
transcribes past the potential termi-nation site.
How does the system ensure that ribosomes initiate
translation as soon as the leader has been synthesized? If ribosomes do not
promptly initiate translation of the leader, premature termination at the
attenu-ator will result. At the typical rates of ribosome binding to messenger,
transcription could easily extend beyond the termination site before the first
ribosome had a chance to bind to messenger and affect loop formation. The
problem of forcing a ribosome onto the mRNA just as it emerges from the
polymerase is solved in a simple way. The trp
leader sequence possesses regions at which transcription is slowed due to
pausing by polymerase. Most likely these pauses result from hairpin structures
in the newly synthesized RNA. The duration of the pauses could be random, but
if the average is long enough, most messengers could then have a ribosome bind
and initiate translation. When polym-erase resumes transcription, the ribosome
following immediately be-hind is properly situated to regulate attenuation.
Related Topics