Chapter: Medical Physiology: The Eye: III. Central Neurophysiology of Vision
Visual Pathways
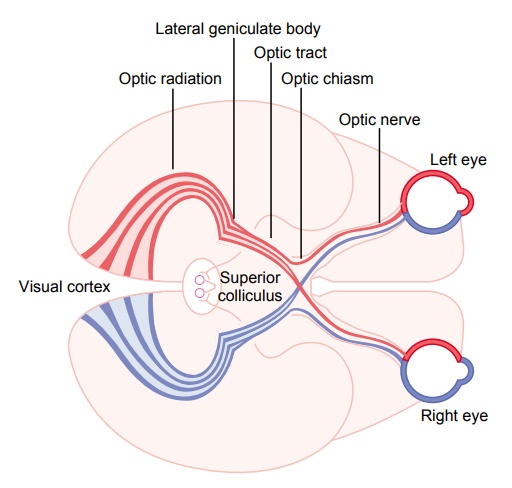
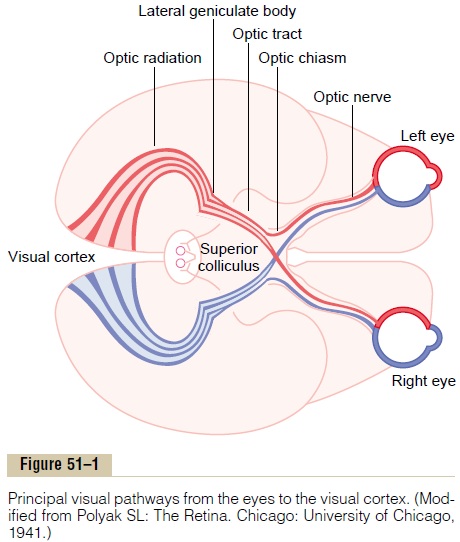
Visual Pathways
Figure 51–1 shows the principal visual pathways from the two retinas to the visual cortex. The visual nerve signals leave the retinas through the opticnerves. At the optic chiasm, the optic nerve fibersfrom the nasal halves of the retinas cross to the opposite sides, where they join the fibers from the opposite temporal retinas to form the optic tracts. The fibers of each optic tract then synapse in the dorsal lateral geniculate nucleus of the thalamus, and from there, geniculocalcarine fibers pass by way of the optic radiation (also called the geniculocalcarine tract) to the primary visual cortex in the calcarine fissure areaof the medial occipital lobe.
Visual fibers also pass to several older areas of the brain: (1) from the optic tracts to the suprachiasmatic nucleus of the hypothalamus, presumably to control circadian rhythms that synchronize various physiologic changes of the body with night and day; (2) into the pretectal nuclei in the midbrain, to elicit reflex movements of the eyes to focus on objects of importance and to activate the pupillary light reflex; (3) into the superior colliculus, to control rapid direc-tional movements of the two eyes; and (4) into the ventral lateral geniculatenucleus of the thalamus and surrounding basal regions of the brain, presumablyto help control some of the body’s behavioral functions.
Thus, the visual pathways can be divided roughly into an old system to the midbrain and base of the forebrain and a new system for direct trans-mission of visual signals into the visual cortex located in the occipital lobes. In human beings, the new system is responsible for perception of virtually all aspects of visual form, colors, and other conscious vision. Conversely, in many primitive animals, even visual form is detected by the older system, using the superior colliculus in the same manner that the visual cortex is used in mammals.
Function of the Dorsal Lateral Geniculate Nucleus of the Thalamus
The optic nerve fibers of the new visual system terminate in the dorsal lateralgeniculate nucleus, located at the dorsal end of the thalamus and also calledsimply the lateral geniculate body, as shown in Figure 51–1. The dorsal lateral geniculate nucleus serves two principal functions: First, it relays visual infor-mation from the optic tract to thevisual cortex by way of the optic radiation (also called the geniculocalcarine tract). This relay function is so accurate that there is exact point-to-point transmission with a high degree of spatial fidelity all the way from the retina to the visual cortex.
It will be recalled that half the fibers in each optic tract after passing the optic chiasm are derived from one eye and half from the other eye, representing cor-responding points on the two retinas. However, the signals from the two eyes are kept apart in the dorsal lateral geniculate nucleus. This nucleus is composed of six nuclear layers. Layers II, III, and V (from ventral to dorsal) receive signals from the lateral half of the ipsilateral retina, whereas layers I, IV, and VI receive signals from the medial half of the retina of the opposite eye. The respective retinal areas of the two eyes connect with neurons that are superimposed over one another in the paired layers, and similar parallel transmission is preserved all the way to the visual cortex.
The second major function of the dorsal lateral geniculate nucleus is to “gate” the transmission of signals to the visual cortex—that is, to control how much of the signal is allowed to pass to the cortex. The nucleus receives gating control signals from two major sources: (1) corticofugal fibers returning in a backward direction from the primary visual cortex to the lateral geniculate nucleus, and (2) reticular areas of the mes-encephalon. Both of these are inhibitory and, whenstimulated, can turn off transmission through selected portions of the dorsal lateral geniculate nucleus. It is assumed that both of these gating circuits help high-light the visual information that is allowed to pass.
Finally, the dorsal lateral geniculate nucleus is divided in another way: (1) Layers I and II are called magnocellular layers because they contain largeneurons.These receive their input almost entirely from the large type Y retinal ganglion cells. This magnocel-lular system provides a rapidly conducting pathway to the visual cortex. However, this system is color blind, transmitting only black-and-white information. Also, its point-to-point transmission is poor because there are not many Y ganglion cells, and their dendrites spread widely in the retina. (2) Layers III through VI are called parvocellular layers because they contain large numbers of small to medium-sized neurons. These neurons receive their input almost entirely from the type X retinal ganglion cells that transmit color and convey accurate point-to-point spatial informa-tion, but at only a moderate velocity of conduction rather than at high velocity.
Related Topics