Chapter: Medical Physiology: The Eye: III. Central Neurophysiology of Vision
Eye Movements and Their Control
Eye Movements and Their Control
To make full use of the visual abilities of the eyes, almost equally as important as interpretation of the visual signals from the eyes is the cerebral control system for directing the eyes toward the object to be viewed.
Muscular Control of Eye Movements. The eye movementsare controlled by three pairs of muscles, shown in Figure 51–7: (1) the medial and lateral recti, (2) the superior and inferior recti, and (3) the superior and inferior obliques. The medial and lateral recti contractto move the eyes from side to side. The superior and inferior recti contract to move the eyes upward or downward. The oblique muscles function mainly to rotate the eyeballs to keep the visual fields in the upright position.
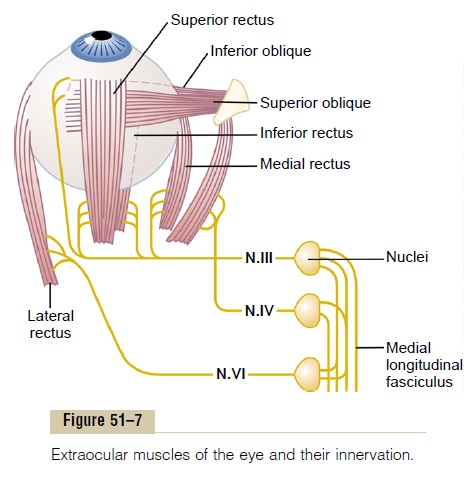
Neural Pathways for Control of Eye Movements. Figure 51–7also shows brain stem nuclei for the third, fourth, and sixth cranial nerves and their connections with the peripheral nerves to the ocular muscles. Shown, too, are interconnections among the brain stem nuclei by way of the nerve tract called the medial longitudinalfasciculus. Each of the three sets of muscles to eacheye is reciprocally innervated so that one muscle of the pair relaxes while the other contracts.
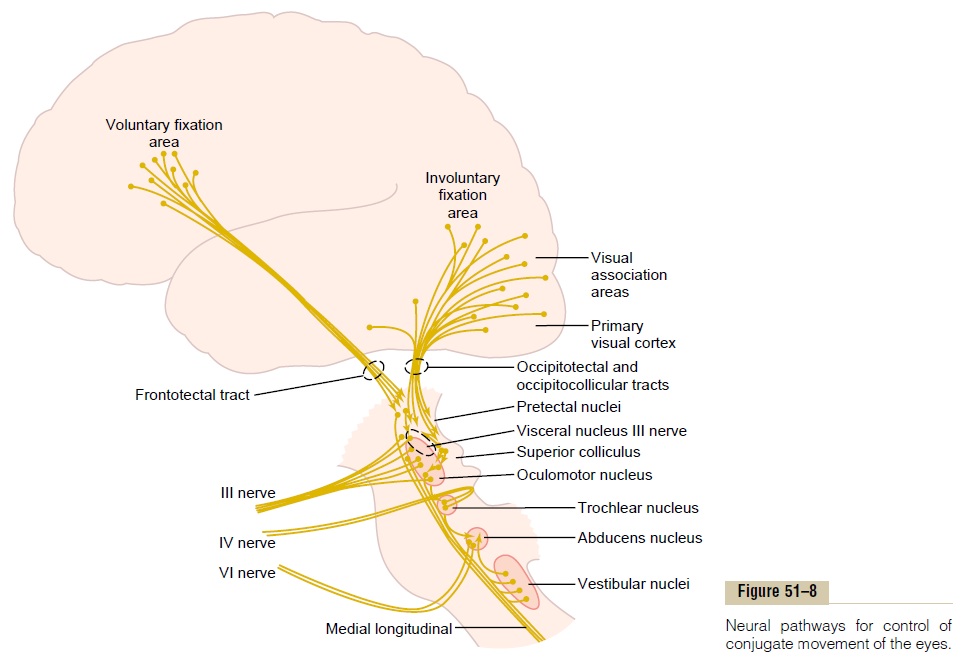
Figure 51–8 demonstrates cortical control of the oculomotor apparatus, showing spread of signals from visual areas in the occipital cortex through occipito-tectal and occipitocollicular tracts to the pretectal and superior colliculus areas of the brain stem. From both the pretectal and the superior colliculus areas, the ocu-lomotor control signals pass to the brain stem nuclei of the oculomotor nerves. Strong signals are also trans-mitted from the body’s equilibrium control centers in the brain stem into the oculomotor system (from the vestibular nuclei by way of the medial longitudinal fasciculus).
Fixation Movements of the Eyes
Perhaps the most important movements of the eyes are those that cause the eyes to “fix” on a discrete portion of the field of vision. Fixation movements are controlled by two neuronal mechanisms. The first of these allows a person to move the eyes voluntarily to find the object on which he or she wants to fix the vision; this is called thevoluntary fixation mechanism.
The second is an involuntary mechanism that holds the eyes firmly on the object once it has been found; this is called the involuntary fixation mechanism.
The voluntary fixation movements are controlled by a cortical field located bilaterally in the premotor cor-tical regions of the frontal lobes, as shown in Figure 51–8. Bilateral dysfunction or destruction of these areas makes it difficult or almost impossible for a person to “unlock” the eyes from one point of fixation and move them to another point. It is usually neces-sary to blink the eyes or put a hand over the eyes for a short time, which then allows the eyes to be moved.
Conversely, the fixation mechanism that causes the eyes to “lock” on the object of attention once it is found is controlled by secondary visual areas in theoccipital cortex,located mainly anterior to the primaryvisual cortex. When this fixation area is destroyed bilaterally in an animal, the animal has difficulty keeping its eyes directed toward a given fixation point or may become totally unable to do so.
To summarize, posterior “involuntary” occipital cor-tical eye fields automatically “lock” the eyes on a given spot of the visual field and thereby prevent movement of the image across the retinas. To unlock this visual fixation, voluntary signals must be transmitted from cortical “voluntary” eye fields located in the frontal cortices.
Mechanism of Involuntary Locking Fixation—Role of the Supe-rior Colliculi. The involuntary locking type of fixationdiscussed in the previous section results from a nega-tive feedback mechanism that prevents the object of attention from leaving the foveal portion of the retina. The eyes normally have three types of continuous but almost imperceptible movements: (1) a continuoustremor at a rate of 30 to 80 cycles per second causedby successive contractions of the motor units in the ocular muscles, (2) aslow drift of the eyeballs in one direction or another, and (3) sudden flicking move-ments that are controlled by the involuntary fixationmechanism.
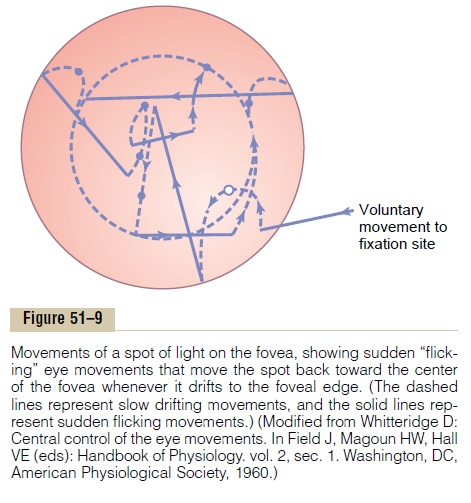
When a spot of light has become fixed on the foveal region of the retina, the tremulous movements cause the spot to move back and forth at a rapid rate across the cones, and the drifting movements cause the spot to drift slowly across the cones. Each time the spot drifts as far as the edge of the fovea, a sudden reflex reaction occurs, producing a flicking movement that moves the spot away from this edge back toward the center of the fovea.Thus, an automatic response moves the image back toward the central point of vision.
These drifting and flicking motions are demon-strated in Figure 51–9, which shows by the dashed lines the slow drifting across the fovea and by the solid lines the flicks that keep the image from leaving the foveal region. This involuntary fixation capability is mostly lost when the superior colliculi are destroyed.
Saccadic Movement of the Eyes—A Mechanism of Successive Fixation Points. When a visual scene is moving contin-ually before the eyes, such as when a person is riding in a car, the eyes fix on one highlight after another in the visual field, jumping from one to the next at a rate of two to three jumps per second. The jumps are called saccades, and the movements are called opticokinetic movements. The saccades occur so rapidly that nomore than 10 per cent of the total time is spent in moving the eyes, with 90 per cent of the time being allocated to the fixation sites. Also, the brain sup-presses the visual image during saccades, so that the person is not conscious of the movements from point to point.
Saccadic Movements During Reading. During theprocess of reading, a person usually makes several sac-cadic movements of the eyes for each line. In this case, the visual scene is not moving past the eyes, but the eyes are trained to move by means of several succes-sive saccades across the visual scene to extract the important information. Similar saccades occur when a person observes a painting, except that the saccades occur in upward, sideways, downward, and angulated directions one after another from one highlight of the painting to another, and so forth.
Fixation on Moving Objects—“Pursuit Movement.” The eyescan also remain fixed on a moving object, which is called pursuit movement. A highly developed cortical mechanism automatically detects the course of move-ment of an object and then rapidly develops a similar course of movement for the eyes. For instance, if an object is moving up and down in a wavelike form at a rate of several times per second, the eyes at first may be unable to fixate on it. However, after a second or so, the eyes begin to jump by means of saccades in approximately the same wavelike pattern of move-ment as that of the object. Then, after another few seconds, the eyes develop progressively smoother movements and finally follow the wave movement almost exactly. This represents a high degree of auto-matic subconscious computational ability by the pursuit system for controlling eye movements.
Superior Colliculi Are Mainly Responsible for Turning the Eyes and Head Toward a Visual Disturbance
Even after the visual cortex has been destroyed, a sudden visual disturbance in a lateral area of the visual field often causes immediate turning of the eyes in that direction. This does not occur if the superior colliculi have also been destroyed. To support this function, the various points of the retina are represented topo-graphically in the superior colliculi in the same way as in the primary visual cortex, although with less accu-racy. Even so, the principal direction of a flash of light in a peripheral retinal field is mapped by the colliculi, and secondary signals are transmitted to the oculo-motor nuclei to turn the eyes. To help in this directional movement of the eyes, the superior colli-culi also have topological maps of somatic sensations from the body and acoustic signals from the ears.
The optic nerve fibers from the eyes to the colliculi that are responsible for these rapid turning move-ments are branches from the rapidly conducting Y fibers, with one branch going to the visual cortex and the other going to the superior colliculi. (The superior colliculi and other regions of the brain stem are also strongly supplied with visual signals transmitted in type W optic nerve fibers. These represent the oldest visual pathway, but their function is unclear.)
In addition to causing the eyes to turn toward a visual disturbance, signals are relayed from the supe-rior colliculi through the medial longitudinal fascicu-lus to other levels of the brain stem to cause turningof the whole head and even of the whole body toward the direction of the disturbance. Other types of non-visual disturbances, such as strong sounds or even stroking of the side of the body, cause similar turning of the eyes, head, and body, but only if the superior col-liculi are intact. Therefore, the superior colliculi play a global role in orienting the eyes, head, and body with respect to external disturbances, whether they are visual, auditory, or somatic.
“Fusion” of the Visual Images from the Two Eyes
To make the visual perceptions more meaningful, the visual images in the two eyes normally fuse with each other on “corresponding points” of the two retinas. The visual cortex plays an important role in fusion. It was pointed out earlier that correspon-ding points of the two retinas transmit visual signals to different neuronal layers of the lateral geniculate body, and these signals in turn are relayed to parallel neurons in the visual cortex. Interactions occur between these cortical neurons to causeinterferenceexcitation in specific neurons when the two visualimages are not “in register”—that is, are not precisely “fused.” This excitation presumably provides the signal that is transmitted to the oculomotor apparatus to cause convergence or divergence or rotation of the eyes so that fusion can be re-established. Once the cor-responding points of the two retinas are in register, excitation of the specific “interference” neurons in the visual cortex disappears.
Neural Mechanism of Stereopsis for Judging Distances of Visual Objects It is pointed out that because the two eyes are more than 2 inches apart, the images on the two retinas are not exactly the same. That is, the right eye sees a little more of the right-hand side of the object, and the left eye a little more of the left-hand side, and the closer the object, the greater the dispar-ity. Therefore, even when the two eyes are fused with each other, it is still impossible for all corresponding points in the two visual images to be exactly in regis-ter at the same time. Furthermore, the nearer the object is to the eyes, the less the degree of register. This degree of nonregister provides the neural mechanism for stereopsis, an important mechanism for judging the distances of visual objects up to about 200 feet (60 meters).
The neuronal cellular mechanism for stereopsis is based on the fact that some of the fiber pathways from the retinas to the visual cortex stray 1 to 2 degrees on each side of the central pathway. Therefore, some optic pathways from the two eyes are exactly in register for objects 2 meters away; still another set of pathways is in register for objects 25 meters away. Thus, the dis-tance is determined by which set or sets of pathways are excited by nonregister or register. This phenome-non is called depth perception, which is another name for stereopsis.
Strabismus
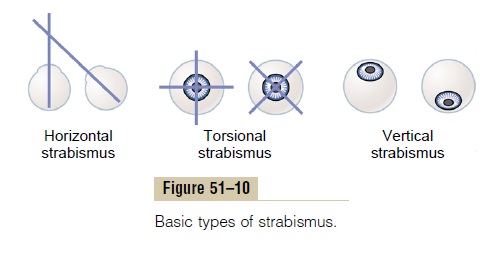
Strabismus, also called squint or cross-eye, means lack of fusion of the eyes in one or more of the visual coor-dinates: horizontal, vertical, or rotational. The basic types of strabismus are shown in Figure 51–10: (1) hor-izontal strabismus, (2) torsional strabismus, and (3) ver-tical strabismus. Combinations of two or even all three of the different types of strabismus often occur.
Strabismus is often caused by abnormal “set” of the fusion mechanism of the visual system. That is, in a young child’s early efforts to fixate the two eyes on the same object, one of the eyes fixates satisfactorily while the other fails do so, or they both fixate satisfactorily but never simultaneously. Soon the patterns of conjugate movements of the eyes become abnormally “set” in the neuronal control pathways themselves, so that the eyes never fuse.
Suppression of the Visual Image from a Repressed Eye. In a fewpatients with strabismus, the eyes alternate in fixing on the object of attention. In other patients, one eye alone is used all the time, and the other eye becomes repressed and is never used for precise vision. The visual acuity of the repressed eye develops only slightly, sometimes remaining 20/400 or less. If the dominant eye then becomes blinded, vision in the repressed eye can develop only to a slight extent in adults but far more in young children. This demonstrates that visual acuity is highly dependent on proper development of central nervous system synaptic connections from the eyes. In fact, even anatomically, the numbers of neuronal con-nections diminish in the visual cortex areas that would normally receive signals from the repressed eye.
Related Topics