Chapter: Medical Physiology: Introduction to Physiology: Genetic Control of Protein Synthesis, Cell Function, and Cell Reproduction
The DNA-Genetic System Also Controls Cell Reproduction
The DNA-Genetic System Also Controls Cell Reproduction
Cell reproduction is another example of the ubiquitous role that the DNA-genetic system plays in all life processes. The genes and their regulatory mechanisms determine the growth characteristics of the cells and also when or whether these cells will divide to form new cells. In this way, the all-important genetic system controls each stage in the development of the human being, from the single-cell fertilized ovum to the whole functioning body. Thus, if there is any central theme to life, it is the DNA-genetic system.
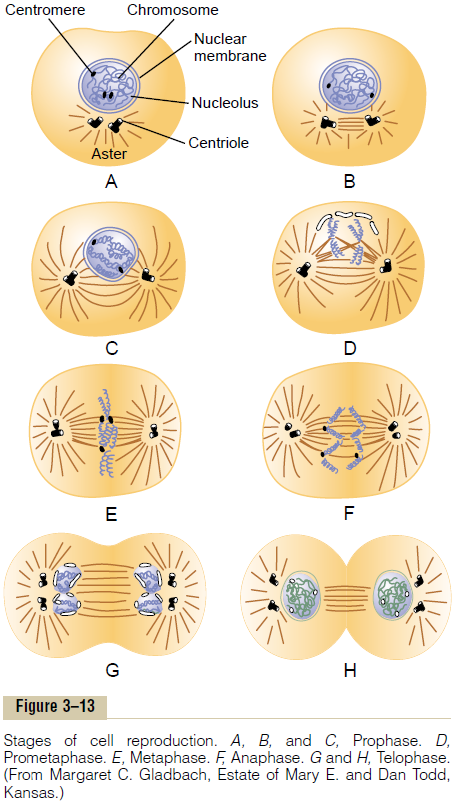
Life Cycle of the Cell. The life cycle of a cell is the periodfrom cell reproduction to the next cell reproduction. When mammalian cells are not inhibited and are repro-ducing as rapidly as they can, this life cycle may be aslittle as 10 to 30 hours. It is terminated by a series of distinct physical events calledmitosis that cause divi-sion of the cell into two new daughter cells. The events of mitosis are shown in Figure 3–13 and are described later. The actual stage of mitosis, however, lasts for only about 30 minutes, so that more than 95 per cent of the life cycle of even rapidly reproducing cells is represented by the interval between mitosis, called interphase.
Except in special conditions of rapid cellular repro-duction, inhibitory factors almost always slow or stop the uninhibited life cycle of the cell. Therefore, differ-ent cells of the body actually have life cycle periods that vary from as little as 10 hours for highly stimu-lated bone marrow cells to an entire lifetime of the human body for most nerve cells.
Cell Reproduction Begins with Replication of DNA
As is true of almost all other important events in the cell, reproduction begins in the nucleus itself. The first step is replication (duplication) of all DNA in the chro-mosomes. Only after this has occurred can mitosis takeplace.
The DNA begins to be duplicated some 5 to 10 hours before mitosis, and this is completed in 4 to 8 hours. The net result is two exact replicas of all DNA. These replicas become the DNA in the two new daughter cells that will be formed at mitosis. After replication of the DNA, there is another period of 1 to 2 hours before mitosis begins abruptly. Even during this period, preliminary changes are beginning to take place that will lead to the mitotic process.
Chemical and Physical Events of DNA Replication. DNA isreplicated in much the same way that RNA is tran-scribed in response to DNA, except for a few impor-tant differences:
1. Both strands of the DNA in each chromosome are replicated, not simply one of them.
2. Both entire strands of the DNA helix are replicated from end to end, rather than small portions of them, as occurs in the transcription of RNA.
3. The principal enzymes for replicating DNA are a complex of multiple enzymes called DNApolymerase, which is comparable to RNApolymerase. It attaches to and moves along the DNA template strand while another enzyme, DNA ligase, causes bonding of successive DNAnucleotides to one another, using high-energy phosphate bonds to energize these attachments.
4. Formation of each new DNA strand occurs simultaneously in hundreds of segments along each of the two strands of the helix until the entire strand is replicated. Then the ends of the subunits are joined together by the DNA ligase enzyme.
5. Each newly formed strand of DNA remains attached by loose hydrogen bonding to the original DNA strand that was used as its template. Therefore, two DNA helixes are coiled together.
6. Because the DNA helixes in each chromosome are approximately 6 centimeters in length and have millions of helix turns, it would be impossible for the two newly formed DNA helixes to uncoil from each other were it not for some special mechanism. This is achieved by enzymes that periodically cut each helix along its entire length, rotate each segment enough to cause separation, and then resplice the helix. Thus, the two new helixes become uncoiled.
DNA Repair, DNA “Proofreading,” and “Mutation.” Duringthe hour or so between DNA replication and the beginning of mitosis, there is a period of very active repair and “proofreading” of the DNA strands. That is, wherever inappropriate DNA nucleotides have been matched up with the nucleotides of the original tem-plate strand, special enzymes cut out the defective areas and replace these with appropriate complemen-tary nucleotides. This is achieved by the same DNA polymerases and DNA ligases that are used in repli-cation. This repair process is referred to as DNA proofreading.
Because of repair and proofreading, the transcrip-tion process rarely makes a mistake. But when a mistake is made, this is called a mutation. The muta-tion causes formation of some abnormal protein in the cell rather than a needed protein, often leading to abnormal cellular function and sometimes even cell death. Yet, given that there are 30,000 or more genes in the human genome and that the period from one human generation to another is about 30 years, one would expect as many as 10 or many more mutations in the passage of the genome from parent to child. As a further protection, however, each human genome is represented by two separate sets of chromosomes with almost identical genes. Therefore, one functional gene of each pair is almost always available to the child despite mutations.
Chromosomes and Their Replication
The DNA helixes of the nucleus are packaged in chro-mosomes. The human cell contains 46 chromosomes arranged in 23 pairs. Most of the genes in the two chro-mosomes of each pair are identical or almost identical to each other, so it is usually stated that the different genes also exist in pairs, although occasionally this is not the case.
In addition to DNA in the chromosome, there is a large amount of protein in the chromosome, composed mainly of many small molecules of electropositively charged histones. The histones are organized into vast numbers of small, bobbin-like cores. Small segments of each DNA helix are coiled sequentially around one core after another.
The histone cores play an important role in the reg-ulation of DNA activity because as long as the DNA is packaged tightly, it cannot function as a template for either the formation of RNA or the replication of new DNA. Further, some of the regulatory proteins have been shown to decondense the histone packaging of the DNA and to allow small segments at a time to form RNA.
Several nonhistone proteins are also major compo-nents of chromosomes, functioning both as chromoso-mal structural proteins and, in connection with the genetic regulatory machinery, as activators, inhibitors, and enzymes.
Replication of the chromosomes in their entirety occurs during the next few minutes after replication of the DNA helixes has been completed; the new DNA helixes collect new protein molecules as needed. The two newly formed chromosomes remain attached to each other (until time for mitosis) at a point called the centromere located near their center. These duplicatedbut still attached chromosomes are called chromatids.
Cell Mitosis
The actual process by which the cell splits into two new cells is called mitosis. Once each chromosome has been replicated to form the two chromatids, in many cells, mitosis follows automatically within 1 or 2 hours.
Mitotic Apparatus: Function of the Centrioles.
One of the first events of mitosis takes place in the cytoplasm, occurring during the latter part of interphase in or around the small structures calledcentrioles. As shown in Figure 3–13, two pairs of centrioles lie close to each other near one pole of the nucleus. (These centrioles, like the DNA and chromosomes, were also replicated during interphase, usually shortly before replication of the DNA.) Each centriole is a small cylindrical body about 0.4 micrometer long and about 0.15 micrometer in diameter, consisting mainly of nine parallel tubular structures arranged in the form of a cylinder. The two centrioles of each pair lie at right angles to each other. Each pair of centrioles, along with attached pericen-triolar material, is called a centrosome.
Shortly before mitosis is to take place, the two pairs of centrioles begin to move apart from each other. This is caused by polymerization of protein microtubules growing between the respective centriole pairs and actually pushing them apart. At the same time, other microtubules grow radially away from each of the cen-triole pairs, forming a spiny star, called the aster, in each end of the cell. Some of the spines of the aster penetrate the nuclear membrane and help separate the two sets of chromatids during mitosis. The complex of microtubules extending between the two new cen-triole pairs is called the spindle, and the entire set of microtubules plus the two pairs of centrioles is called the mitotic apparatus.
Prophase. The first stage of mitosis, calledprophase,isshown in Figure 3–13A, B, and C. While the spindle is forming, the chromosomes of the nucleus (which in interphase consist of loosely coiled strands) become condensed into well-defined chromosomes.
Prometaphase. During this stage (see Figure 3–13D),the growing microtubular spines of the aster fragment the nuclear envelope. At the same time, multiple microtubules from the aster attach to the chromatids at the centromeres, where the paired chromatids are still bound to each other; the tubules then pull one chromatid of each pair toward one cellular pole and its partner toward the opposite pole.
Metaphase. During metaphase (see Figure 3–13E), thetwo asters of the mitotic apparatus are pushed farther apart. This is believed to occur because the micro-tubular spines from the two asters, where they inter-digitate with each other to form the mitotic spindle, actually push each other away. There is reason to believe that minute contractile protein molecules called “motor molecules,” perhaps composed of the muscle protein actin, extend between the respective spines and, using a stepping action as in muscle, actively slide the spines in a reverse direction along each other. Simultaneously, the chromatids are pulled tightly by their attached microtubules to the very center of the cell, lining up to form the equatorial plate of the mitotic spindle.
Anaphase. During this phase (see Figure 3–13F), thetwo chromatids of each chromosome are pulled apart at the centromere. All 46 pairs of chromatids are separated, forming two separate sets of 46 daughter chromosomes. One of these sets is pulled toward onemitotic aster and the other toward the other aster as the two respective poles of the dividing cell are pushed still farther apart.
Telophase. In telophase (see Figure 3–13GandH), thetwo sets of daughter chromosomes are pushed com-pletely apart. Then the mitotic apparatus dissolutes, and a new nuclear membrane develops around each set of chromosomes. This membrane is formed from portions of the endoplasmic reticulum that are already present in the cytoplasm. Shortly thereafter, the cell pinches in two, midway between the two nuclei. This is caused by formation of a contractile ring of micro-filaments composed of actin and probably myosin (thetwo contractile proteins of muscle) at the juncture of the newly developing cells that pinches them off from each other.
Control of Cell Growth and Cell Reproduction
We know that certain cells grow and reproduce all the time, such as the blood-forming cells of the bone marrow, the germinal layers of the skin, and the epithe-lium of the gut. Many other cells, however, such as smooth muscle cells, may not reproduce for many years. A few cells, such as the neurons and most stri-ated muscle cells, do not reproduce during the entire life of a person, except during the original period of fetal life.
In certain tissues, an insufficiency of some types of cells causes these to grow and reproduce rapidly until appropriate numbers of them are again available. For instance, in some young animals, seven eighths of the liver can be removed surgically, and the cells of the remaining one eighth will grow and divide until the liver mass returns almost to normal. The same occurs for many glandular cells and most cells of the bone marrow, subcutaneous tissue, intestinal epithelium, and almost any other tissue except highly differenti-ated cells such as nerve and muscle cells.
We know little about the mechanisms that maintain proper numbers of the different types of cells in the body. However, experiments have shown at least three ways in which growth can be controlled. First, growth often is controlled by growth factors that come from other parts of the body. Some of these circulate in the blood, but others originate in adjacent tissues. For instance, the epithelial cells of some glands, such as the pancreas, fail to grow without a growth factor from the sublying connective tissue of the gland. Second, most normal cells stop growing when they have run out of space for growth. This occurs when cells are grown in tissue culture; the cells grow until they contact a solid object, and then growth stops. Third, cells grown in tissue culture often stop growing when minute amounts of their own secretions are allowed to collect in the culture medium. This, too, could provide a means for negative feedback control of growth.
Regulation of Cell Size. Cell size is determined almostentirely by the amount of functioning DNA in the nucleus. If replication of the DNA does not occur, the cell grows to a certain size and thereafter remains at that size. Conversely, it is possible, by use of the chem-ical colchicine, to prevent formation of the mitotic spindle and therefore to prevent mitosis, even though replication of the DNA continues. In this event, the nucleus contains far greater quantities of DNA than it normally does, and the cell grows proportionately larger. It is assumed that this results simply from increased production of RNA and cell proteins, which in turn cause the cell to grow larger.
Related Topics