Chapter: Medical Physiology: Introduction to Physiology: Genetic Control of Protein Synthesis, Cell Function, and Cell Reproduction
DNA Code in the Cell Nucleus Is Transferred to an RNA Code in the Cell Cytoplasm - The Process of Transcription
The DNA Code in the Cell Nucleus Is Transferred to an RNA Code in the Cell Cytoplasm - The Process of Transcription
Because the DNA is located in the nucleus of the cell, yet most of the functions of the cell are carried out in the cytoplasm, there must be some means for the DNA genes of the nucleus to control the chemical reactions of the cytoplasm. This is achieved through the inter-mediary of another type of nucleic acid, RNA, the for-mation of which is controlled by the DNA of the nucleus. Thus, as shown in Figure 3–7, the code is trans-ferred to the RNA; this process is called transcription.
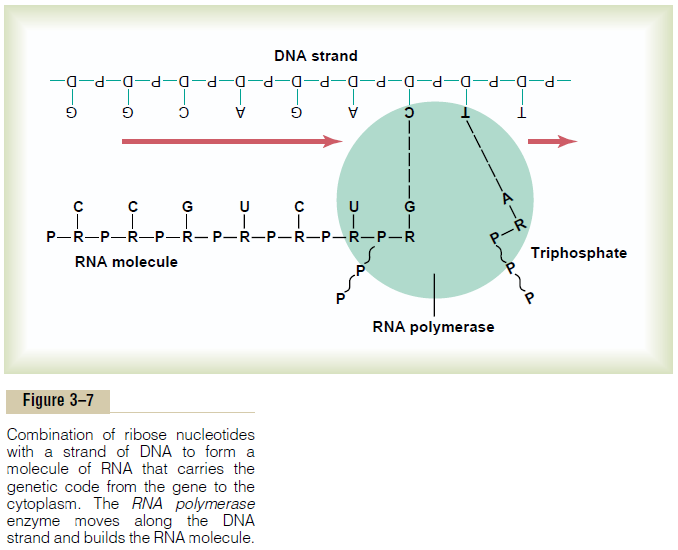
The RNA, in turn, diffuses from the nucleus through nuclear pores into the cytoplasmic compartment, where it controls protein synthesis.
Synthesis of RNA
During synthesis of RNA, the two strands of the DNA molecule separate temporarily; one of these strands is used as a template for synthesis of an RNA molecule. The code triplets in the DNA cause formation of com-plementary code triplets (called codons) in the RNA;these codons, in turn, will control the sequence of amino acids in a protein to be synthesized in the cell cytoplasm.
Basic Building Blocks of RNA. The basic building blocks ofRNA are almost the same as those of DNA, except for two differences. First, the sugar deoxyribose is not used in the formation of RNA. In its place is another sugar of slightly different composition, ribose, containing an extra hydroxyl ion appended to the ribose ring struc-ture. Second, thymine is replaced by another pyrimi-dine, uracil.
Formation of RNA Nucleotides. The basic building blocksof RNA form RNA nucleotides, exactly as previously described for DNA synthesis. Here again, four sepa-rate nucleotides are used in the formation of RNA. These nucleotides contain the bases adenine, guanine,cytosine, and uracil. Note that these are the same bases as in DNA, except that uracil in RNA replaces thymine in DNA.
“Activation” of the RNA Nucleotides. The next step inthe synthesis of RNA is “activation” of the RNA nucleotides by an enzyme, RNA polymerase. This occurs by adding to each nucleotide two extra phos-phate radicals to form triphosphates (shown in Figure 3–7 by the two RNA nucleotides to the far right during RNA chain formation). These last two phosphates are combined with the nucleotide by high-energy phos-phate bonds derived from ATP in the cell.
The result of this activation process is that large quantities of ATP energy are made available to each of the nucleotides, and this energy is used to promote the chemical reactions that add each new RNA nucleotide at the end of the developing RNA chain.
Assembly of the RNA Chain from Activated Nucleotides Using the DNA Strand as a Template—The Process of “Transcription”
Assembly of the RNA molecule is accomplished in the manner shown in Figure 3–7 under the influence of an enzyme, RNA polymerase. This is a large protein enzyme that has many functional properties necessary for formation of the RNA molecule. They are as follows:
1. In the DNA strand immediately ahead of the initial gene is a sequence of nucleotides called the promoter. The RNA polymerase has an appropriate complementary structure that recognizes this promoter and becomes attached to it. This is the essential step for initiating formation of the RNA molecule.
2. After the RNA polymerase attaches to the promoter, the polymerase causes unwinding of about two turns of the DNA helix and separation of the unwound portions of the two strands.
3. Then the polymerase moves along the DNA strand, temporarily unwinding and separating the two DNA strands at each stage of its movement. As it moves along, it adds at each stage a new activated RNA nucleotide to the end of the newly forming RNA chain by the following steps:
a. First, it causes a hydrogen bond to form between the end base of the DNA strand and the base of an RNA nucleotide in the nucleoplasm.
b. Then, one at a time, the RNA polymerase breaks two of the three phosphate radicals away from each of these RNA nucleotides, liberating large amounts of energy from the broken high-energy phosphate bonds; this energy is used to cause covalent linkage of the remaining phosphate on the nucleotide with the ribose on the end of the growing RNA chain.
c. When the RNA polymerase reaches the end of the DNA gene, it encounters a new sequence of DNA nucleotides called the chain-terminating sequence; this causes the polymerase and the newly formed RNA chain to break away from the DNA strand. Then the polymerase can be used again and again to form still more new RNA chains.
d. As the new RNA strand is formed, its weak hydrogen bonds with the DNA template break away, because the DNA has a high affinity for rebonding with its own complementary DNA strand. Thus, the RNA chain is forced away from the DNA and is released into the nucleoplasm.
Thus, the code that is present in the DNA strand is eventually transmitted in complementary form to the RNA chain. The ribose nucleotide bases always combine with the deoxyribose bases in the following combinations:
Three Different Types of RNA. There are three differenttypes of RNA, each of which plays an independent and entirely different role in protein formation:
1. Messenger RNA, which carries the genetic code tothe cytoplasm for controlling the type of protein formed.
2. Transfer RNA, which transports activated aminoacids to the ribosomes to be used in assembling the protein molecule.
3. Ribosomal RNA, which, along with about 75different proteins, forms ribosomes, the physical and chemical structures on which protein molecules are actually assembled.
Messenger RNA—The Codons
Messenger RNA molecules are long, single RNAstrands that are suspended in the cytoplasm. These molecules are composed of several hundred to several thousand RNA nucleotides in unpaired strands, and they contain codons that are exactly complementary to the code triplets of the DNA genes. Figure 3–8 shows a small segment of a molecule of messenger
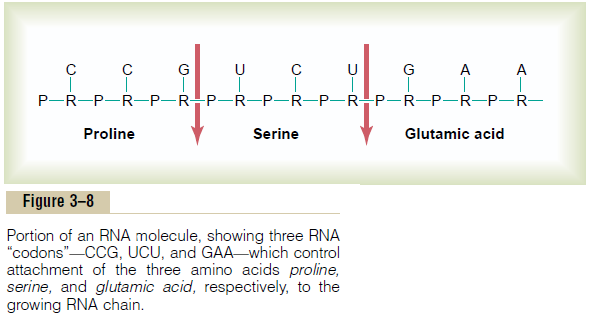
RNA. Its codons are CCG, UCU, and GAA. These are the codons for the amino acids proline, serine, and glu-tamic acid. The transcription of these codons from the DNA molecule to the RNA molecule is shown in Figure 3–7.
RNA Codons for the Different Amino Acids. Table 3–1 givesthe RNA codons for the 20 common amino acids found in protein molecules. Note that most of the amino acids are represented by more than one codon; also, one codon represents the signal “start manufac-turing the protein molecule,” and three codons repre-sent “stop manufacturing the protein molecule.” In Table 3–1, these two types of codons are designated CI for “chain-initiating” and CT for “chain-terminating.”

Transfer RNA—The Anticodons
Another type of RNA that plays an essential role in protein synthesis is called transfer RNA, because it transfers amino acid molecules to protein molecules as the protein is being synthesized. Each type of transfer RNA combines specifically with 1 of the 20 amino acids that are to be incorporated into proteins. The transfer RNA then acts as a carrier to transport its specific type of amino acid to the ribosomes, where protein molecules are forming. In the ribosomes, each specific type of transfer RNA recognizes a particular codon on the messenger RNA (described later) and thereby delivers the appropriate amino acid to the appropriate place in the chain of the newly forming protein molecule.
Transfer RNA, which contains only about 80 nucleotides, is a relatively small molecule in com-parison with messenger RNA. It is a folded chain of nucleotides with a cloverleaf appearance similar to that shown in Figure 3–9. At one end of the molecule is always an adenylic acid; it is to this that the trans-ported amino acid attaches at a hydroxyl group of the ribose in the adenylic acid.
Because the function of transfer RNA is to cause attachment of a specific amino acid to a forming protein chain, it is essential that each type of transfer RNA also have specificity for a particular codon in the

messenger RNA. The specific code in the transfer RNA that allows it to recognize a specific codon is again a triplet of nucleotide bases and is called ananti-codon. This is located approximately in the middleof the transfer RNA molecule (at the bottom of the cloverleaf configuration shown in Figure 3–9). During formation of the protein molecule, the anticodon bases combine loosely by hydrogen bonding with the codon bases of the messenger RNA. In this way, the respec-tive amino acids are lined up one after another along the messenger RNA chain, thus establishing the appropriate sequence of amino acids in the newly forming protein molecule.
Ribosomal RNA
The third type of RNA in the cell is ribosomal RNA; it constitutes about 60 per cent of the ribosome. The remainder of the ribosome is protein, containing about 75 types of proteins that are both structural proteins and enzymes needed in the manufacture of protein molecules.
The ribosome is the physical structure in the cyto-plasm on which protein molecules are actually syn-thesized. However, it always functions in association with the other two types of RNA as well: transfer RNA transports amino acids to the ribosome for incorpora-tion into the developing protein molecule, whereas messenger RNA provides the information necessaryfor sequencing the amino acids in proper order for each specific type of protein to be manufactured.
Thus, the ribosome acts as a manufacturing plant in which the protein molecules are formed.
Formation of Ribosomes in the Nucleolus. The DNA genesfor formation of ribosomal RNA are located in five pairs of chromosomes in the nucleus, and each of these chromosomes contains many duplicates of these par-ticular genes because of the large amounts of riboso-mal RNA required for cellular function.
As the ribosomal RNA forms, it collects in the nucleolus, a specialized structure lying adjacent to thechromosomes.When large amounts of ribosomal RNA are being synthesized, as occurs in cells that manufac-ture large amounts of protein, the nucleolus is a large structure, whereas in cells that synthesize little protein, the nucleolus may not even be seen. Ribosomal RNA is specially processed in the nucleolus, where it binds with “ribosomal proteins” to form granular condensa-tion products that are primordial subunits of ribo-somes. These subunits are then released from the nucleolus and transported through the large pores of the nuclear envelope to almost all parts of the cyto-plasm. After the subunits enter the cytoplasm, they are assembled to form mature, functional ribosomes. Therefore, proteins are formed in the cytoplasm of the cell, but not in the cell nucleus, because the nucleus does not contain mature ribosomes.
Formation of Proteins on the Ribosomes—The Process of “Translation”
When a molecule of messenger RNA comes in contact with a ribosome, it travels through the ribosome, beginning at a predetermined end of the RNA mole-cule specified by an appropriate sequence of RNA bases called the “chain-initiating” codon. Then, as shown in Figure 3–9, while the messenger RNA travels through the ribosome, a protein molecule is formed— a process called translation. Thus, the ribosome reads the codons of the messenger RNA in much the same way that a tape is “read” as it passes through the play-back head of a tape recorder. Then, when a “stop” (or “chain-terminating”) codon slips past the ribosome, the end of a protein molecule is signaled and the protein molecule is freed into the cytoplasm.
Polyribosomes. A single messenger RNA molecule canform protein molecules in several ribosomes at the same time because the initial end of the RNA strand can pass to a successive ribosome as it leaves the first, as shown at the bottom left in Figure 3–9 and in Figure 3–10. The protein molecules are in different stages of development in each ribosome. As a result, clusters of ribosomes frequently occur, 3 to 10 ribosomes being attached to a single messenger RNA at the same time. These clusters are called polyribosomes.

It is especially important to note that a messenger RNA can cause the formation of a protein molecule in any ribosome; that is, there is no specificity of ribo-somes for given types of protein. The ribosome is simply the physical manufacturing plant in which the chemical reactions take place.
Many Ribosomes Attach to the Endoplasmic Reticulum. It was noted that many ribosomes become attached to the endoplasmic reticulum. This occurs because the initial ends of many forming protein mol-ecules have amino acid sequences that immediately attach to specific receptor sites on the endoplasmic reticulum; this causes these molecules to penetrate the reticulum wall and enter the endoplasmic reticulum matrix. This gives a granular appearance to those por-tions of the reticulum where proteins are being formed and entering the matrix of the reticulum.
Figure 3–10 shows the functional relation of mes-senger RNA to the ribosomes and the manner in which the ribosomes attach to the membrane of the endoplasmic reticulum. Note the process of translation occurring in several ribosomes at the same time in response to the same strand of messenger RNA. Note also the newly forming polypeptide (protein) chains passing through the endoplasmic reticulum membrane into the endoplasmic matrix.
Yet it should be noted that except in glandular cells in which large amounts of protein-containing secre-tory vesicles are formed, most proteins synthesized by the ribosomes are released directly into the cytosol instead of into the endoplasmic reticulum. These pro-teins are enzymes and internal structural proteins of the cell.
Chemical Steps in Protein Synthesis. Some of the chemicalevents that occur in synthesis of a protein molecule are shown in Figure 3–11. This figure shows representative reactions for three separate amino acids, AA1, AA2, and AA20.

The stages of the reactions are the follow-ing: (1) Each amino acid is activated by a chemical process in which ATP combines with the amino acid to form an adenosine monophosphate complex with theamino acid, giving up two high-energy phosphatebonds in the process. (2) The activated amino acid, having an excess of energy, then combines with itsspecific transfer RNA to form an amino acid–tRNA complex and, at the same time, releases the adenosinemonophosphate. (3) The transfer RNA carrying the amino acid complex then comes in contact with the messenger RNA molecule in the ribosome, where the anticodon of the transfer RNA attaches tem-porarily to its specific codon of the messenger RNA, thus lining up the amino acid in appropriate sequence to form a protein molecule. Then, under the influence of the enzyme peptidyl transferase (one of the proteins in the ribosome), peptide bonds are formed between the successive amino acids, thus adding progressively to the protein chain. These chemical events require energy from two additional high-energy phosphate bonds, making a total of four high-energy bonds used for each amino acid added to the protein chain. Thus, the synthesis of proteins is one of the most energy-con-suming processes of the cell.
Peptide Linkage. The successive amino acids in theprotein chain combine with one another according to the typical reaction:

In this chemical reaction, a hydroxyl radical (OH–) is removed from the COOH portion of the first amino acid, and a hydrogen (H+) of the NH2portion of the other amino acid is removed. These combine to form water, and the two reactive sites left on the two suc-cessive amino acids bond with each other, resulting in a single molecule. This process is called peptidelinkage. As each additional amino acid is added, anadditional peptide linkage is formed.
Related Topics