Chapter: Plant Biochemistry: Polysaccharides are storage and transport forms of carbohydrates produced by photosynthesis
Surplus of photosynthesis products can be stored temporarily in chloroplasts as starch
Surplus of photosynthesis products can be stored temporarily in chloroplasts as starch
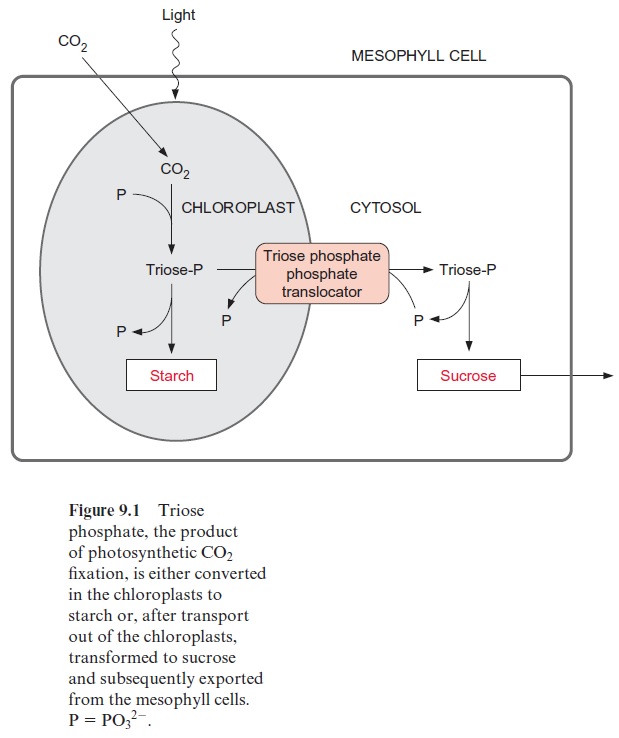
Figure 9.12 outlines the synthesis and degradation of transitory starch in chloroplasts. The regulation of ADP-glucose pyrophosphorylase by 3-phosphoglycerate (3-PGA) and phosphate (P) enables the control of the flux of carbohydrates into starch. The activity of the enzyme is governed by the 3-PGA/P concentration ratio. 3-PGA is a major metabolite in the chloroplast stroma. Due to the equilibrium of the reactions catalyzed by phosphoglycerate kinase and glyceraldehyde phosphate dehydrogenase , the stromal 3-PGA concentration is much higher than that of triose phosphate. In the chloroplast stroma the total amount of phos-phate and phosphorylated intermediates of the Calvin cycle is virtually kept constant by the counter-exchange of the triose phosphate-phosphate translocator . Therefore, an increase of the 3-PGA concentra-tion results in a decrease of the phosphate concentration. The 3-PGA/P ratio is therefore a very sensitive indicator of the metabolite status in the chloroplast stroma. When sucrose synthesis is decreased, leading to a decrease of phosphate release in the cytosol, the chloroplasts would suf-fer from phosphate deficiency, which would limit photosynthesis in the chloroplasts (Fig. 9.1). Under such conditions, however, the PGA/P quo-tient increases, which enhances starch synthesis and the resulting release of phosphate allows photosynthesis to continue. In this case starch acts as a buffer. Assimilates that are not utilized for synthesis of sucrose or other metabolites are deposited temporarily in the chloroplasts as transi-tory starch. Moreover, starch synthesis is programmed in such a way (by a mechanism largely unknown) that sufficient starch is deposited each day for use during the following night.
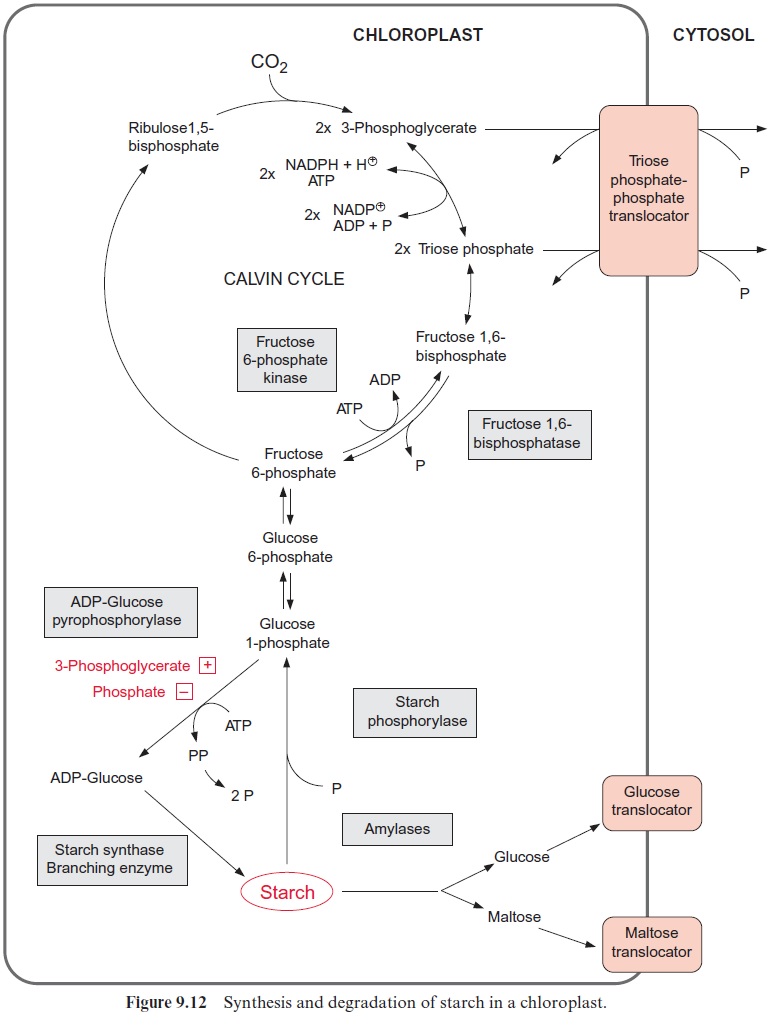
So far very little is known about the regulation of transitory starch deg-radation. It probably is stimulated by an increase in the stromal phosphate concentration, but the mechanism for this is still unclear. An increase in the stromal phosphate concentration indicates a shortage of substrates. Hydrolytic starch degradation leads to the release of maltose and glucose, which are transported into the cytosol via specific translocators. Maltose, frequently the main product of starch degradation, is split in the cytosol by atransglucosidase. This enzyme transfers one glucose molecule of mal-tose to an -glucan present in the cytosol with the release of the remain-ing glucose molecule, which is phosphorylated by ATP via a hexokinase to glucose 6-phosphate. The cytosolic -glucan is degraded by a cytosolic phosphorylase.
Glucose 1-phosphate, derived from phosphorolytic starch degradation, is converted in a reversal of the starch synthesis pathway to fructose 6-phosphate and the latter to fructose 1,6-bisphosphate by fructose 6-phosphate kinase. Triose phosphate formed from fructose 1,6-bisphosphate by aldolase is released from the chloroplasts via the triose phosphate-phosphate trans-locator. Part of the triose phosphate is oxidized within the chloroplasts to 3-phosphoglycerate and is subsequently exported also via the triose phos-phate-phosphate translocator. This translocator as well as the glucose translocator is thus involved in the mobilization of the chloroplast transitory starch.
Related Topics