Chapter: Biotechnology Applying the Genetic Revolution: Basics of biotechnology
Subviral Infectious Agents and Other Gene Creatures
SUBVIRAL
INFECTIOUS AGENTS AND OTHER GENE CREATURES
We have used the term gene creatures to refer to various genetic entities that are sometimes called subviral infectious agents. These creatures exist, but are not considered living because none of them can produce their own energy, duplicate their own genomes, or live independent of a host. The main advantage a virus has over a gene creature is the ability to survive as an inactive particle outside the host cell. Gene creatures are not normally found outside the host cell.
Satellite viruses are defective viruses. They can either
replicate their genome or package their genome
into a capsid, but they are unable to do both by themselves. Satellite viruses
rely on a
helper virus to supply the
missing components or genes. For example, hepatitis delta virus (HDV) is a
small single-stranded RNA satellite virus that infects the liver. Its helper is
hepatitis B virus. Bacteriophage P4 is a satellite virus that infects E. coli.
It is a double-stranded DNA
virus that can replicate as a plasmid or integrate into the host chromosome,
but it cannot form virus particles by itself. It relies on P2 bacteriophage
to supply the structural proteins. P4
sends transcription factors to the P2 genome to control expression of the genes
it pirates.
Gene creatures also include
genetic elements that may be helpful to the host. For example, the plasmids of
E. coli and yeast are genetic elements that cannot produce their own energy and
rely on the host cell to replicate their genome. They cannot survive outside a
host cell. These traits qualify plasmids as gene creatures. Like viruses and
satellite viruses, plasmids are replicons, that is, they have sufficient
information in their genome to direct their own replication.
Plasmids may confer positive
traits to the host. For example, plasmids can provide antibacterial enzymes,
such as bacteriocins, that help their host compete with other bacteria for
nutrients (see earlier discussion). Plasmids may carry genes for antibiotic
resistance, thus allowing the host bacteria
to survive after encountering an antibiotic. Plasmids may confer
virulence, making the host bacteria more aggressive and deadly. Finally, some
plasmids contain genes that help the host degrade a new carbon source to
provide food. Plasmids are usually found as circles of DNA, although some
linear plasmids have been found. Plasmids come in all sizes, but are usually
much smaller than the bacterial chromosome. The genes on plasmids are often
beneficial to the host.
Because the plasmid coexists
within the cytoplasm of the host cell, it does not generally harm its host.
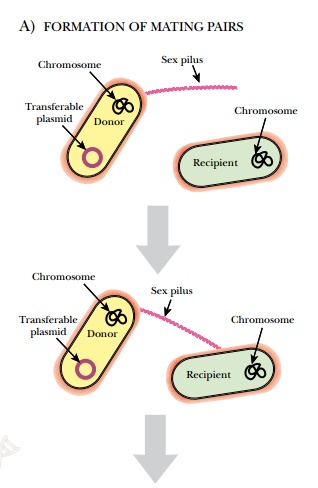
The F plasmid is found in some
E. coli, and it is about 1% of the size of the chromosome. It was named “F” for
fertility because it confers the ability to mate. F plasmids can transfer
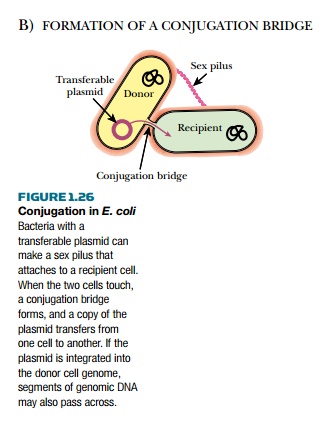
themselves from one cell to the next in a process called conjugation (Fig.
1.26). The plasmid has genes for the formation of a specialized pilus, the
F-pilus, which physically attaches an F+E. coli to an F− cell. After contact, a
Conjugation in E. coli junction—the
conjugation bridge—forms between the two cells. During replication of the F
plasmid, one strand is cut at the origin and the free end enters the cytoplasm
of the F− cell via the conjugation bridge. Inside the recipient a complementary
strand of DNA is made and the plasmid is recircularized. The other strand of
the parent plasmid remains in the original F+ cell and is also duplicated.
Thus, after conjugation, both cells become F+. Occasionally, the F plasmid
integrates into the host chromosome. If an integrated F plasmid is transferred
to another cell via conjugation, parts of the host chromosome may also get
transferred.
Therefore, bacteria can
exchange chromosomal genetic information through conjugation. Another gene
creature that is very useful in biotechnology is the transposable element or
transposon. This genetic element is merely a length of DNA that cannot exist or
replicate as an independent molecule. To survive, it integrates into another
DNA molecule. Mobile DNA or jumping genes are two terms used to describe
transposons. When the transposon moves from one location to another, the process
is called transposition. Unlike plasmids,
transposons lack an origin of
replication and are not considered replicons. They can only be replicated by
integrating themselves into a host DNA molecule, such as a chromosome, plasmid,
or viral genome. Transposons can move from site to site within the same host
DNA or move from one host molecule of DNA to another. If a transposon loses its
ability to move, its DNA remains in place on the chromosome or other DNA
molecule. This is what comprises much human “junk DNA”.
Transposons come in several
varieties and are classified based on the mechanism of movement. Transposons
have two inverted DNA repeats at each end and a gene for transposase, the enzyme needed for movement. Transposase recognizes
the inverted repeats at the ends of
the transposon and excises
the entire element from the chromosome. Next, transposase recognizes a target sequence of 3 to 9 base pairs in
length on the host DNA. The transposon is then inserted into the target
sequence, which is duplicated in the process. One copy is found on each side of
the transposon. When a transposon is completely removed from one site and moved
to another, the mechanism is conservative
transposition or cut-and-paste transposition (Fig. 1.27). This leaves
behind a double-stranded break that must be repaired by the host cell. Several cellular mechanisms exist to make this
type of repair.
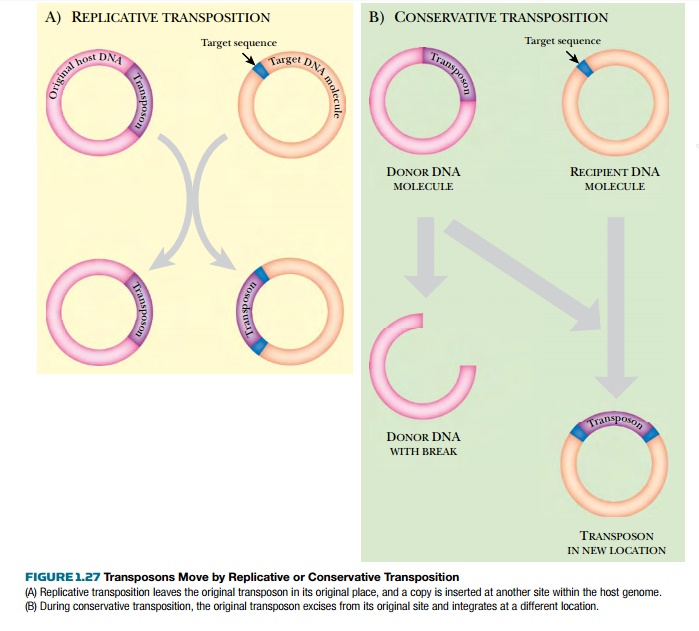
An alternative mechanism is replicative transposition, where a
second copy of the transposon is made. Complex
transposons use this method. Much as before, transposase recognizes the
inverted repeats of the transposon. However, in this case it only makes
single-stranded nicks at the ends. Transposase then makes two single-stranded
nicks, one at each end of the target site. Each single DNA strand of the
transposon is joined to one host strand at the target site. This creates two
single-stranded copies of the transposon. The host responds to such
single-stranded DNA regions by making the second, complementary strand of the
transposon. This gives two copies of the transposon. Notice how the transposon
itself does not replicate. It tricks the host into making the replica.
Transposon movement can cause
problems for the host. When the transposon moves, there is a potential for
insertions, deletions, and inversions in the host DNA. If two copies of a
transposon are found on a plasmid and the target sequence is on the host
chromosome, a segment of the plasmid (flanked by the transposons) may be
inserted into the host DNA. More generally, when multiple transposons are near
each other, the ends of two neighboring but separate transposons may be used
for transposition. When the two ends move to a new location, the DNA between
them will be carried along. Whole genes or segments of genes may be deleted
from the original location in this process. Conversely, regions of chromosome
may become duplicated. If transposons are active and move often, the genome
will become very damaged and the host cells often commit suicide Because the transposon
will be destroyed along with its host, many transposons move only rarely.
Controlling their movement preserves their existence within the genome and
keeps the host cell from committing suicide.
Gene creatures is a term to describe genetic elements that
exist within the confines of a host cell, yet are separate from the original
host genome. Some gene creatures include satellite viruses, plasmids, and
transposons.
The plasmid is a unique gene creature because it confers
positive traits such as resistance to antibiotics, bacteriocins, and the
ability to transfer genetic material between two cells.
Transposons do not contain origins for their independent
replication as do plasmids. These elements subvert the cell to make their
copies by inducing breaks in the genome.
Related Topics