Chapter: Plant Biochemistry: Biotechnology alters plants to meet requirements of agriculture, nutrition and industry
Selected promoters enable the defined expression of a foreign gene
Selected promoters enable the defined expression of a foreign gene
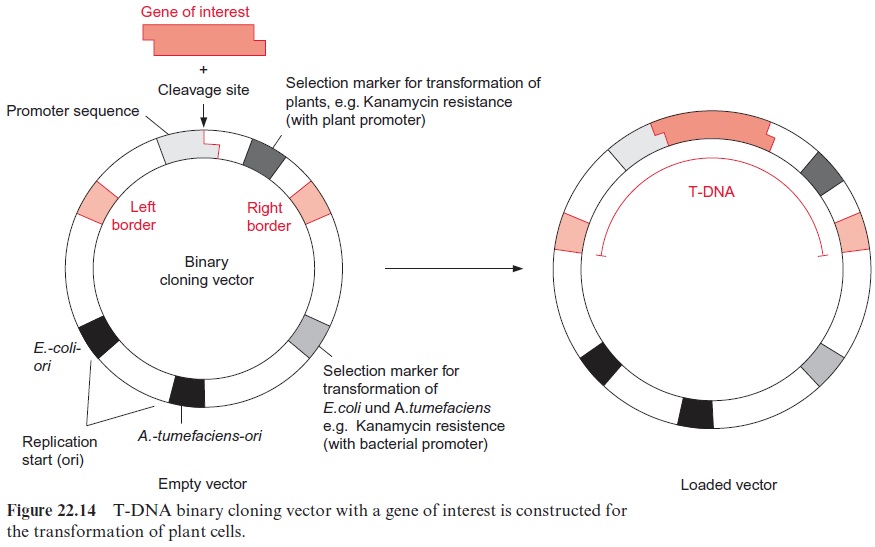
Any foreign gene transferred to a plant can be expressed only when it has been provided with a suitable promoter . The selection of the promoter determines where, when, how much, and under what conditions gene expression takes place. Usually promoters are already included in the commercially available vectors (see Fig. 22.14).
The CaMV-35S promoter from the cauliflower mosaic virus is often used to reach high expression levels of an inserted foreign gene in all parts of the plants. This promoter comprises transcription enhancers that enable a particularly high transcription rate in different tissues of very many plants. The nos promoter from the Ti-plasmid of A. tumefaciens, normally regulating the gene for nopaline synthesis, is also often used as a nonspecific promoter in transgenic plants.
The DNA sequences of specific plant promoters are identified by the analysis of plant genes obtained from a genomic gene library. A promoter for specific gene expression in the potato tuber, for instance, has been identified by analysis of the DNA sequence of the gene for pata-tin, the storage protein of the potato .
A reporter gene can be used to determine whether an isolated promoter sequence is tissue specific. Such reporter genes encode proteins that are easily detected in a plant, for instance, the green fluorescent protein (GFP) from the jellyfishAequorea victoria, which can be seen directly by confocal microscopy in intact leaves. Frequently, the gene for the β-glucuronidase (GUS) enzyme from E. coli also is used as a reporter gene (Fig. 22.19). This enzyme resembles β-galactosidase and does not occur in plants. It hydrolyzes a synthetic X-glucuronide which releases a blue hydrolysis product, easily identifiable under the microscope. When a potato plant is transformed with the GUS reporter gene fused to the pata-tin promoter, after addition of X-glucuronide, a deep blue color develops only in cuts of the tubers, but not in other tissues of the plant. Thus, the patatin promoter acts as an organ-specific promoter. Many promoters have been isolated that are active only in certain organs or tissues, such as leaves, roots, phloem, flowers, or seed. Moreover, promoters have been isolated that control the expression of gene products only under certain environmental conditions, such as light, high temperatures, water stress, or pathogenic infection. Frequently, promoters are also active in heterologous plant species, although the extent of expression by a certain promoter can vary between different plant species.

Gene products are directed to certain subcellular compartments by targeting sequences
In transgenic plants, the expression of a foreign gene can be restricted to certain tissue or cell types by the use of defined promoters. Furthermore, a protein encoded by a foreign gene can be directed to a particular sub-cellular destination (e.g., the chloroplast stroma or the vacuolar compart-ment) by the presence of additional amino acid. Such amino acids can bepresequences or transit peptides, which serve as targeting signals for trans-fer via the various subcellular membrane transport systems. Many targeting sequences are now available, which can be used in gene technology to direct a foreign protein to a defined subcellular com-partment, such as the vacuolar compartment or the chloroplast stroma.
Related Topics