Chapter: Plant Biochemistry: The Calvin cycle catalyzes photosynthetic CO2 assimilation
Ribulose bisphosphate is regenerated from triose phosphate
Ribulose bisphosphate is regenerated from triose phosphate
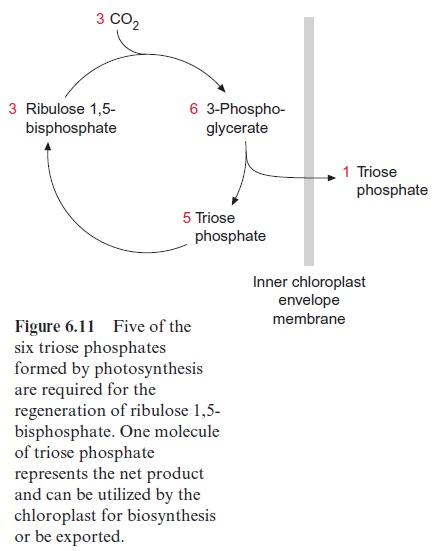
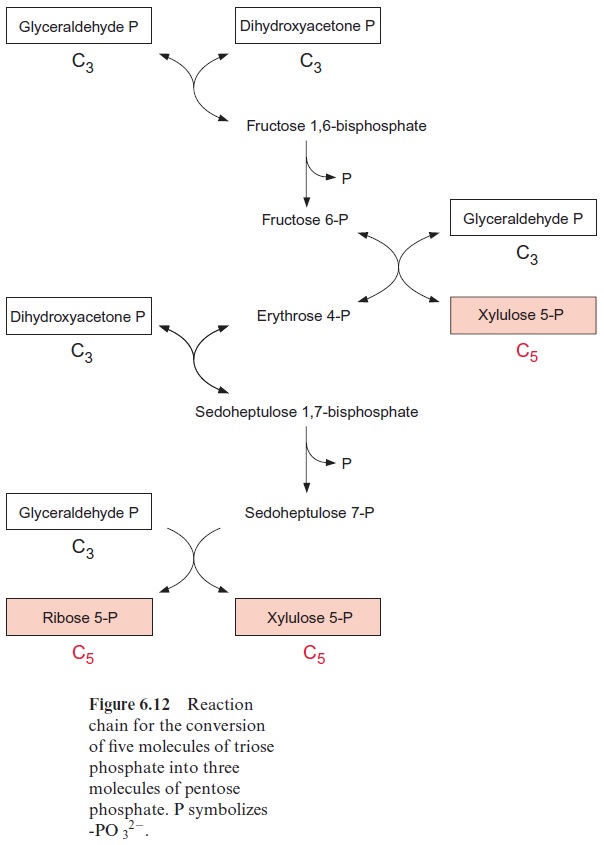
The fixation of three molecules of CO2 in the Calvin cycle results in the synthesis of six molecules of phosphoglycerate which are converted to six molecules of triose phosphate (Fig. 6.11). Of these, only one molecule of triose phosphate is the actual gain, which is provided to the cell for various biosynthetic processes. The remaining five triose phosphates are needed to regenerate three molecules of ribulose bisphosphate so that the Calvin cycle can continue. Figure 6.12 shows the metabolic pathway of the conversion of the five triose phosphates (white boxes) to three pentose phosphates (red boxes).
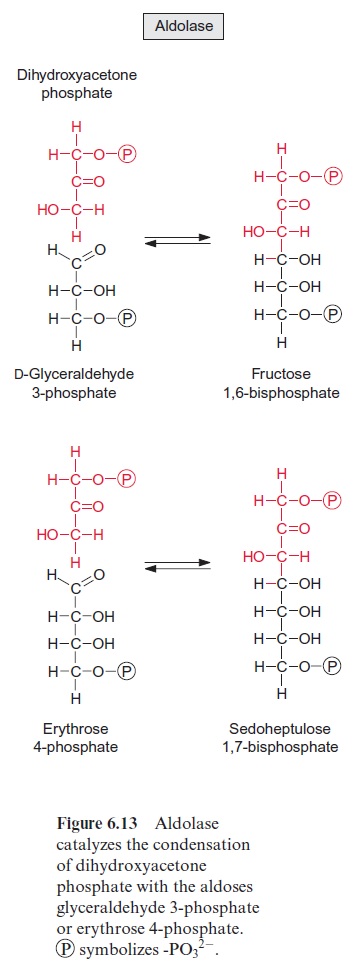
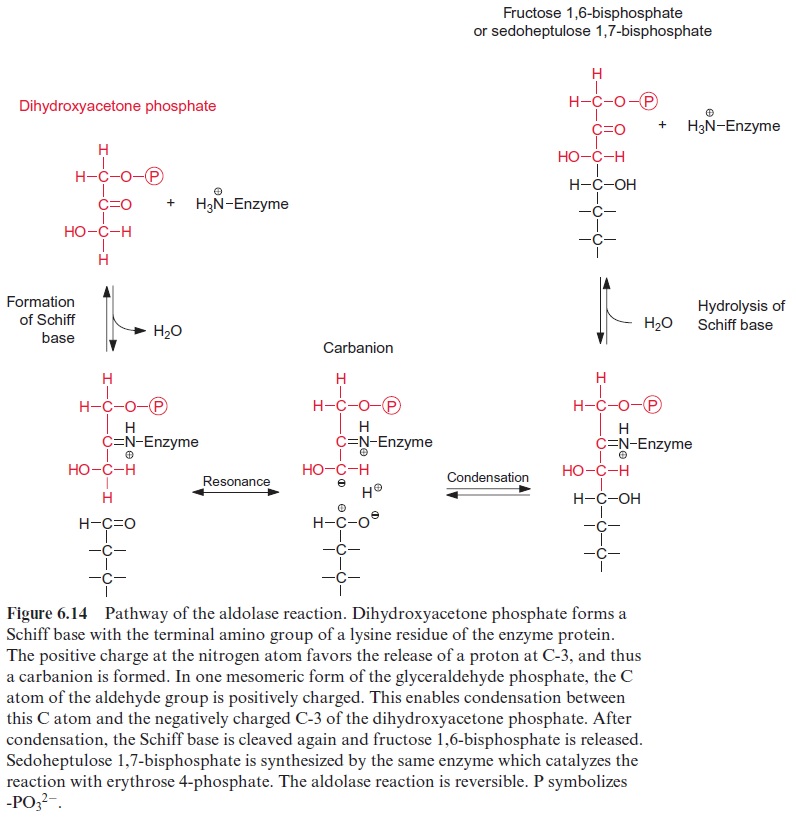
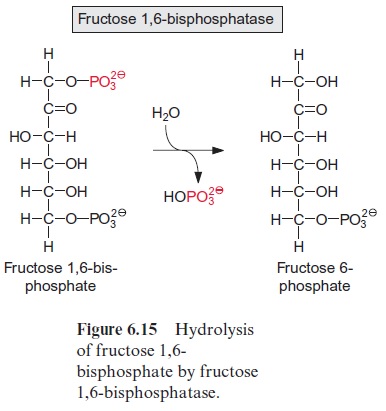
The two trioses dihydroxyacetone phosphate and glyceraldehyde phos phate are condensed in a reversible reaction to fructose 1,6-bisphosphate, as catalyzed by the enzyme aldolase (Fig. 6.13). Figure 6.14 shows the reaction mechanism. As an intermediate of this reaction, a protonated Schiff base is formed between a lysine residue of the active center of the enzyme and the keto group of dihydroxyacetone phosphate. This Schiff base enhances the release of a proton from the C-3 position and enables a nucleophilic reac tion with the C atom of the aldehyde group of glyceraldehyde phosphate. Fructose 1,6-bisphosphate is hydrolyzed by fructose 1,6-bisphosphatase in an irreversible reaction to fructose 6-phosphate (Fig. 6.15).
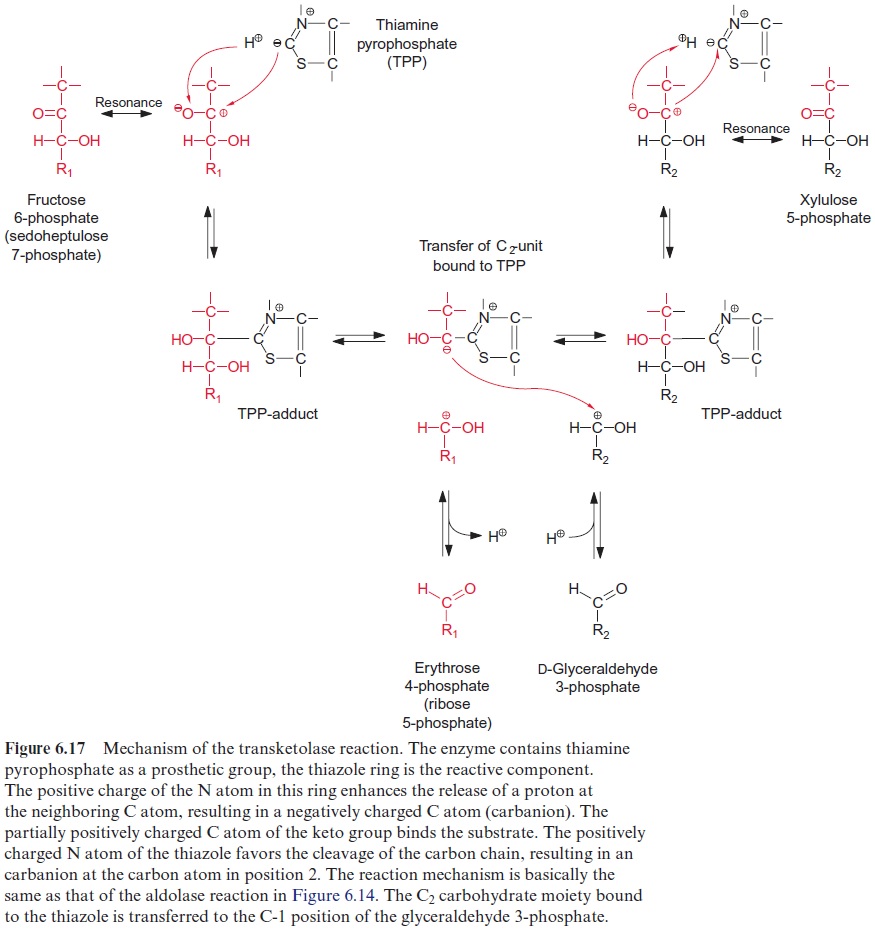
The enzyme transketolase transfers a carbohydrate residue with two carbon atoms from fructose 6-phosphate to glyceraldehyde 3-phosphate yielding xylulose 5-phosphate, and erythrose 4-phosphate in a reversible reaction (Fig. 6.16). Thiamine pyrophosphate (Fig. 5.5), already discussed as a reaction partner of pyruvate oxidation , is involved as the prosthetic group in this reaction (Fig. 6.17).
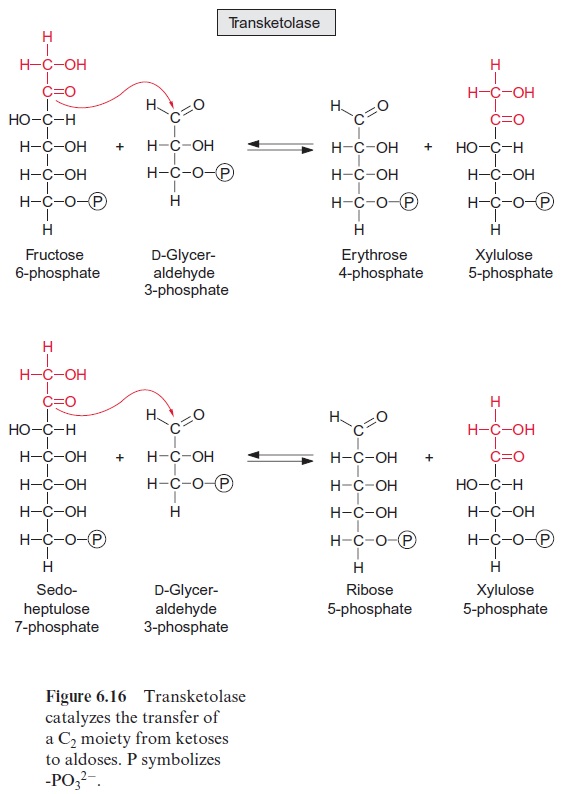
Once more an aldolase (Fig. 6.13) catalyzes a condensation reaction; erythrose 4-phosphate is combined with dihydroxyacetone phosphate to form sedoheptulose 1,7-bisphosphate. Subsequently, the enzyme sedoheptu-lose 1,7-bisphosphatase catalyzes the irreversible hydrolysis of sedoheptulose 1,7-bisphosphate. This reaction is similar to the hydrolysis of fructose 1,6-bisphosphate, despite the fact that both reactions are catalyzed by different enzymes. Again, a carbohydrate residue of two C atoms is transferred bytransketolase from sedoheptulose 7-phosphate to dihydroxyacetone phos phate to form ribose 5-phosphate and xylulose 5-phosphate (Fig. 6.16).
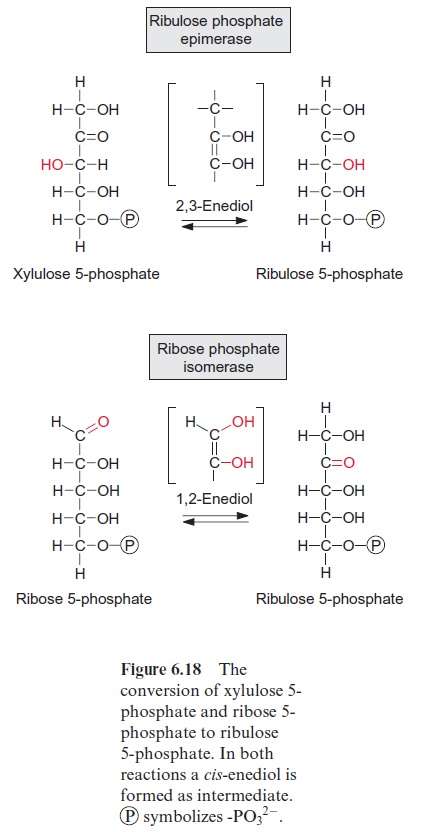
The three pentose phosphates synthesized are then converted to ribu lose 5-phosphate (Fig. 6.18). The conversion of xylulose 5-phosphate is catalyzed by ribulose phosphate epimerase; this reaction proceeds via a keto-enol isomerization and a 2,3-enediol intermediate. The conversion of the aldose ribose 5-phosphate to the ketose ribulose 5-phosphate is cata lyzed by ribose phosphate isomerase, again via an enediol as intermediate, although in the 1,2-position. The three molecules of ribulose 5-phosphate formed in this way are converted to the CO2 acceptor ribulose 1,5-bisphos phate upon consumption of ATP by ribulose phosphate kinase (Fig. 6.19).
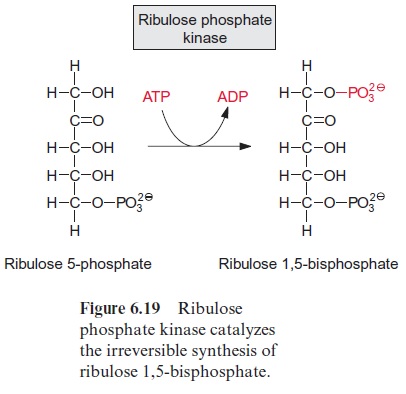
This kinase reaction is irreversible, since a phosphate of the “energy-rich” anhydride in the ATP is converted to a phosphate ester with a low free energy of hydrolysis.
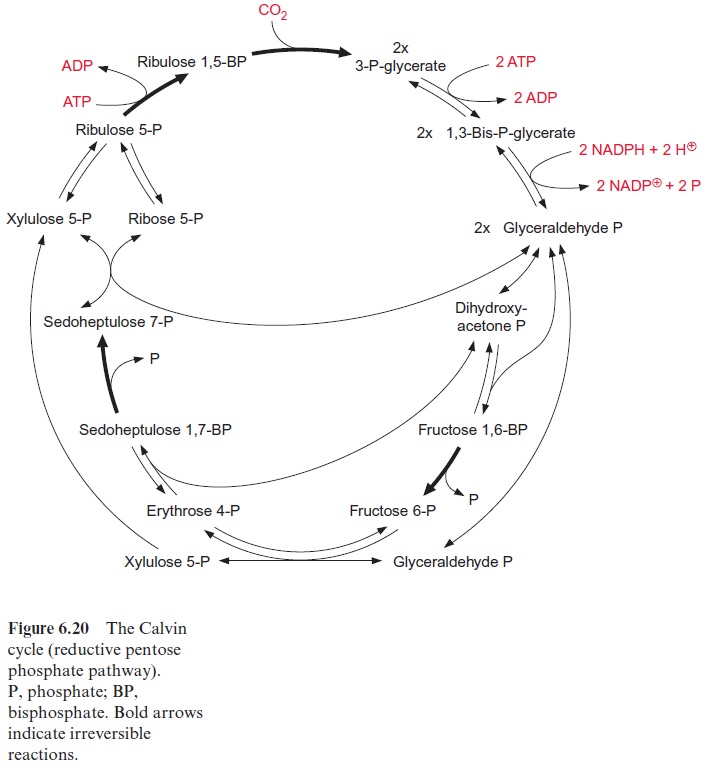
The scheme in Figure 6.20 presents a summary of the various reactions of the Calvin cycle. Four irreversible steps are indicated in the cycle (bold arrows): carboxylation, hydrolysis of fructose- and sedoheptulose bisphosphate,and phosphorylation of ribulose 5-phosphate. The fixation of one molecule of CO2 requires in total two molecules of NADPH and three molecules of ATP.
Related Topics