Chapter: Plant Biology : Floral development and reproductive physiology
Plant Physiology of floral initiation and development
PHYSIOLOGY OF FLORAL INITIATION AND DEVELOPMENT
Key Notes
Floral meristems
Flowers originate from the shoot meristem. The change in the vegetative meristem to develop a flower is termed induction. This is followed by evocation, the development of the floral meristem. The flower then forms and becomes functional when the reproductive structures are mature. Flowers are formed in concentric whorls of sepals, petals, stamens and carpels.
Floral evocation
Floral evocation usually requires an external stimulus such as cold (vernalization) sensed by the meristem, or appropriate daylength sensed by phytochrome, species being long-day, short-day or day-neutral. Hormones are important in the control of flowering.
Floral development genes
Heterochrony (flowering-time) genes regulate the conversion of the vegetative meristem to a floral meristem. Floral meristem identity genes then regulate the formation of a flower. When flowering has been initiated, cadastral genes govern the formation of the whorls. Finally, homeotic genes control the structure of the flower, influenced by the cadastral genes. The ABC model of flower development predicts that the four whorls of the flower are controlled by three homeotic genes A, B and C. Mutations of these genes form flowers in which the organs are misplaced.
Floral meristems
Flowers originate from the shoot meristem which normally generates leaves and shoots. The meristem stops vegetative growth when flowering commences and either produces a single flower (determinate or closed inflorescence) or a succession of floral meristems, each of which will become a flower (indeterminate or open inflorescence). Like a shoot meristem, a floral meristem is divided into layers: the tunica producing the outer cell layers, and the corpus the inner cell layers. The first stage of the flowering process is termed induction, the change in form and function of the vegetative meristem to develop a flower. This is followed by evocation, the development of the floral meristem. The flower itself is then formed and becomes functional when the reproductive structures are mature.
Floral evocation
Forming a flower requires the differentiation of the vegetative shoot meristem into a floral meristem when it has reached an appropriate stage of development. Flowering usually requires one of a range of external stimuli that
bring about floral evocation. In temperate plants, this may be vernalization, a chilling period preceding flowering, possibly by weeks or months, or daylength, species being classified as long-day, short-day or day-neutral species. Vernalization appears to be sensed by the meristem itself, as chilling the plant while the meristem is warmed does not induce flowering. Day length is sensed by phytochrome in the young leaves, suggesting that ahormone is involved in transmitting the signal to the meristem. This flowering hormone was originally named florigen, but its existence has never been proven.
Floral development genes
Genes for many aspects of the process of floral development have been described in two species, arabidopsis and Antirrhinum majus. It will be helpful to have read The arabidopsis flower is actinomorphic and mainly self-pollinated, while the Antirrhinum flower is zygomorphic and insect-pollinated.
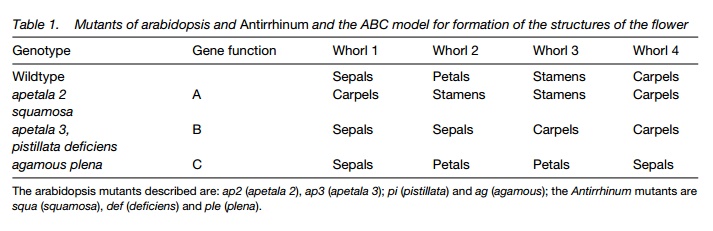
The earliest stages of flower formation involve the activity of genes known as heterochrony or flowering-time genes which regulate the conversion of the vegetative meristem to a floral meristem. Once this has happened, flower meristem identity genes regulate the formation of the flower. The Arabidopsis mutant known as leafy (lfy), for instance, that has a mutation in a flower meristem identity gene, forms shoots where flowers should be. Once flowering has been initiated, a third group of genes known as cadastral genes are initiated which govern the formation of the whorls of the flower. Finally, the structure of the flower is governed by homeotic genes, which cause the right structure to appear in the right place. The function of homeotic genes is influenced by the cadastral genes expressed before them. The ABC model of flower development predicts that the four whorls of the flower are controlled by the action of three genes A, B and C. By studying floral mutants affecting each of these genes (Table1), the way in which they control development has been established.
Each whorl is specified by the activity of one or two of the three homeotic genes, A, B and C, where: A alone ®sepals; A and B ®petals; B and C®stamens; C alone ® carpels. This will occur regardless of where A, B and C are active in the flower; so a mutant, where B is inactive, makes two whorls of sepals and no petals (Table 1). A and C inhibit each other: if A is inactive, C becomes more active and vice versa, so a plant without C will form petals in whorls 2 and 3 and sepals in whorls 1 and 4. Sepals form in whorl 4, because the action of the A gene in this whorl was being inhibited by the activity of the C gene.
Related Topics