Chapter: Medical Physiology: Regulation of Extracellular Fluid Osmolarity and Sodium Concentration
Osmoreceptor-ADH Feedback System
Osmoreceptor-ADH Feedback System
Figure 28–8 shows the basic components of the osmoreceptor-ADH feedback system for control of extracellular fluid sodium concentration and osmolarity. When osmolarity (plasma sodium concentration) increases above normal because of water deficit, for example, this feedback system operates as follows:
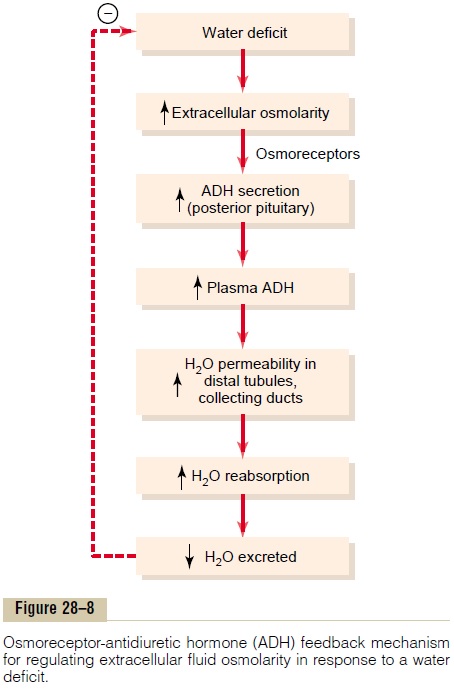
1. An increase in extracellular fluid osmolarity (which in practical terms means an increase in plasma sodium concentration) causes the special nerve cells called osmoreceptor cells, located in the anterior hypothalamus near the supraoptic nuclei, to shrink.
2. Shrinkage of the osmoreceptor cells causes them to fire, sending nerve signals to additional nerve cells in the supraoptic nuclei, which then relay these signals down the stalk of the pituitary gland to the posterior pituitary.
3. These action potentials conducted to the posterior pituitary stimulate the release of ADH, which is stored in secretory granules (or vesicles) in the nerve endings.
4. ADH enters the blood stream and is transported to the kidneys, where it increases the water permeability of the late distal tubules, cortical collecting tubules, and medullary collecting ducts.
5. The increased water permeability in the distal nephron segments causes increased water reabsorption and excretion of a small volume of concentrated urine.
Thus, water is conserved in the body while sodium and other solutes continue to be excreted in the urine. This causes dilution of the solutes in the extracellular fluid, thereby correcting the initial excessively concentrated extracellular fluid.
The opposite sequence of events occurs when the extracellular fluid becomes too dilute (hypo-osmotic). For example, with excess water ingestion and a decrease in extracellular fluid osmolarity, less ADH is formed, the renal tubules decrease their permeability for water, less water is reabsorbed, and a large volume of dilute urine is formed. This in turn concentrates the body fluids and returns plasma osmolarity toward normal.
ADH Synthesis in Supraoptic and Paraventricular Nuclei of the Hypothalamus and ADH Release from the Posterior Pituitary
Figure 28–9 shows the neuroanatomy of the hypothal-amus and the pituitary gland, where ADH is synthe-sized and released. The hypothalamus contains two types of magnocellular (large) neurons that synthesizeADH in the supraoptic and paraventricular nuclei of the hypothalamus, aboutfive sixths in the supraopticnuclei and about one sixth in the paraventricular nuclei. Both of these nuclei have axonal extensions to the posterior pituitary. Once ADH is synthesized, it is transported down the axons of the neurons to their tips, terminating in the posterior pituitary gland. When the supraoptic and paraventricular nuclei are stimu-lated by increased osmolarity or other factors, nerve impulses pass down these nerve endings, changing their membrane permeability and increasing calcium entry. ADH stored in the secretory granules (also called vesicles) of the nerve endings is released in response to increased calcium entry. The released ADH is then carried away in the capillary blood of the posterior pituitary into the systemic circulation.
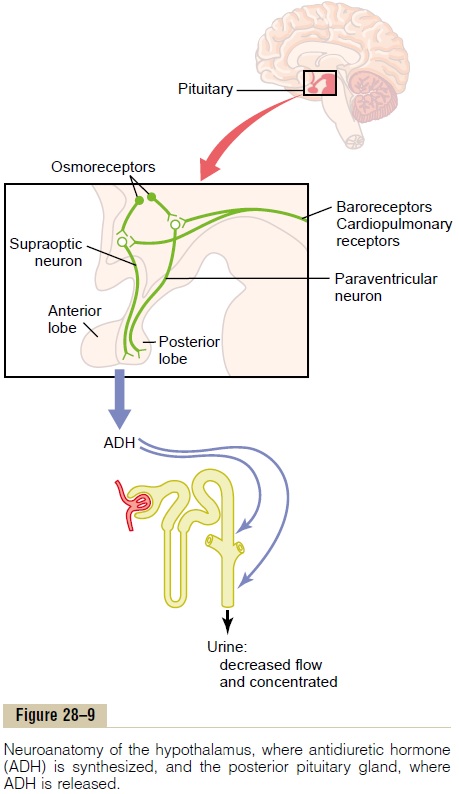
Secretion of ADH in response to an osmotic stimu-lus is rapid, so that plasma ADH levels can increase severalfold within minutes, thereby providing a rapid means for altering renal excretion of water.
A second neuronal area important in controlling osmolarity and ADH secretion is located along the anteroventral region of the third ventricle, called the AV3V region. At the upper part of this region is astructure called the subfornical organ, and at the infe-rior part is another structure called the organum vas-culosum of the lamina terminalis. Between these twoorgans is the median preoptic nucleus, which has mul-tiple nerve connections with the two organs as well as with the supraoptic nuclei and the blood pressure control centers in the medulla of the brain. Lesions of the AV3V region cause multiple deficits in the control of ADH secretion, thirst, sodium appetite, and blood pressure. Electrical stimulation of this region or stim-ulation by angiotensin II can alter ADH secretion, thirst, and sodium appetite.
In the vicinity of the AV3V region and the supraop-tic nuclei are neuronal cells that are excited by small increases in extracellular fluid osmolarity; hence, the term osmoreceptors has been used to describe these neurons. These cells send nerve signals to the supraop-tic nuclei to control their firing and secretion of ADH.
It is also likely that they induce thirst in response to increased extracellular fluid osmolarity.
Both the subfornical organ and the organum vascu-losum of the lamina terminalis have vascular supplies that lack the typical blood-brain barrier that impedes the diffusion of most ions from the blood into the brain tissue. This makes it possible for ions and other solutes to cross between the blood and the local interstitial fluid in this region. As a result, the osmoreceptors rapidly respond to changes in osmolarity of the extra-cellular fluid, exerting powerful control over the secre-tion of ADH and over thirst, as discussed later.
Cardiovascular Reflex Stimulation of ADH Release by Decreased Arterial Pressure and/or Decreased Blood Volume
ADH release is also controlled by cardiovascular reflexes that respond to decreases in blood pressure and/or blood volume, including (1) the arterial barore-ceptor reflexes and (2) the cardiopulmonary reflexes. These reflex pathways originate in high-pressure regions of the cir-culation, such as the aortic arch and carotid sinus, and in the low-pressure regions, especially in the cardiac atria. Afferent stimuli are carried by the vagus and glossopharyngeal nerves with synapses in the nuclei of the tractus solitarius. Projections from these nuclei relay signals to the hypothalamic nuclei that control ADH synthesis and secretion.
Thus, in addition to increased osmolarity, two other stimuli increase ADH secretion: (1) decreased arterial pressure and (2) decreased blood volume. Whenever blood pressure and blood volume are reduced, such as occurs during hemorrhage, increased ADH secretion causes increased fluid reabsorption by the kidneys, helping to restore blood pressure and blood volume toward normal.
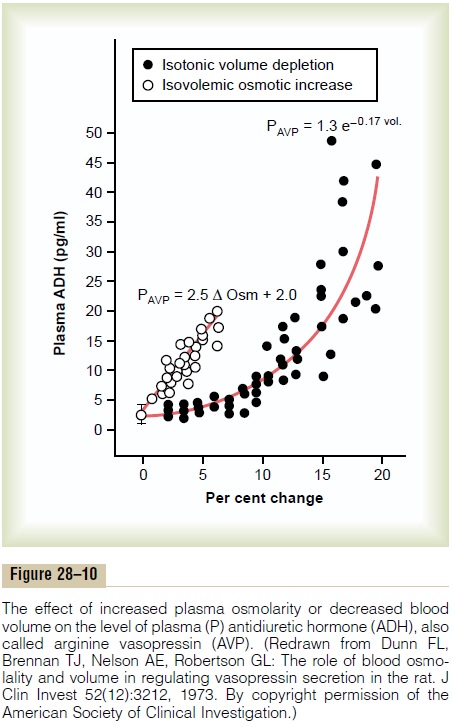
Quantitative Importance of Cardiovascular Reflexes and Osmolarity in Stimulating ADH Secretion
As shown in Figure 28–10, either a decrease in effec-tive blood volume or an increase in extracellular fluid osmolarity stimulates ADH secretion. However, ADH is considerably more sensitive to small changes in osmolarity than to similar changes in blood volume. For example, a change in plasma osmolarity of only 1 per cent is sufficient to increase ADH levels. By contrast, after blood loss, plasma ADH levels do not change appreciably until blood volume is reduced by about 10 per cent. With further decreases in blood volume,ADH levels rapidly increase. Thus, with severe decreases in blood volume, the cardiovascular reflexes play a major role in stimulating ADH secretion. The usual day-to-day regulation of ADH secretion during simple dehydration is effected mainly by changes in plasma osmolarity. Decreased blood volume, however,
Other Stimuli for ADH Secretion
ADH secretion can also be increased or decreased by other stimuli to the central nervous system as well as by various drugs and hormones, as shown in Table 28–2. For example, nausea is a potent stimulus for ADH release, which may increase to as much as 100 times normal after vomiting. Also, drugs such as nico-tine and morphine stimulate ADH release, whereassome drugs, such as alcohol, inhibit ADH release. The marked diuresis that occurs after ingestion of alcohol is due in part to inhibition of ADH release.

Related Topics