Chapter: Medical Physiology: The Eye: II. Receptor and Neural Function of the Retina
Neural Circuitry of the Retina - Neural Function of the Retina
Neural Function of the Retina
Neural Circuitry of the Retina
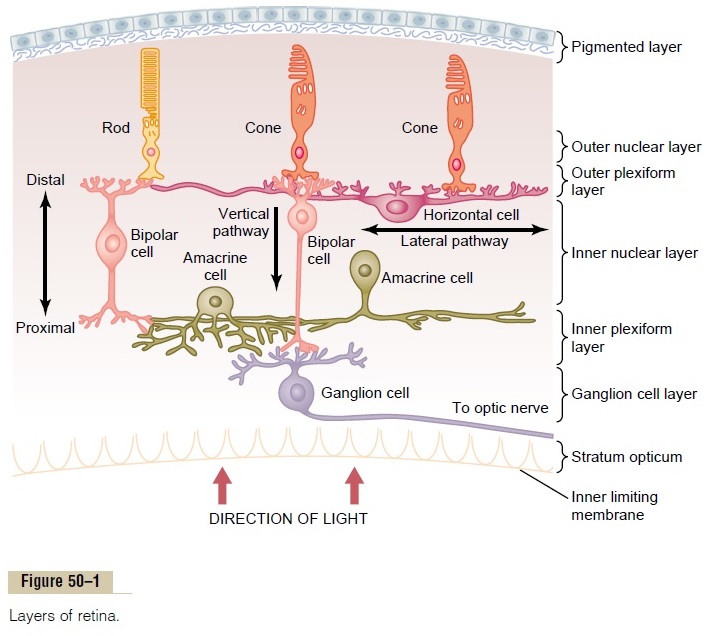
Figure 50–1 shows the tremendous complexity of neural organization in the retina. To simplify this, Figure 50–11 presents the essentials of the retina’s neural connections, showing at the left the circuit in the peripheral retina and at the right the circuit in the foveal retina. The different neuronal cell types are as follows:
1. The photoreceptors themselves—the rods and cones—which transmit signals to the outerplexiform layer, where they synapse with bipolar cells and horizontal cells
2. The horizontal cells, which transmit signals horizontally in the outer plexiform layer from the rods and cones to bipolar cells
3. The bipolar cells, which transmit signals vertically from the rods, cones, and horizontal cells to the inner plexiform layer, where they synapse with ganglion cells and amacrine cells
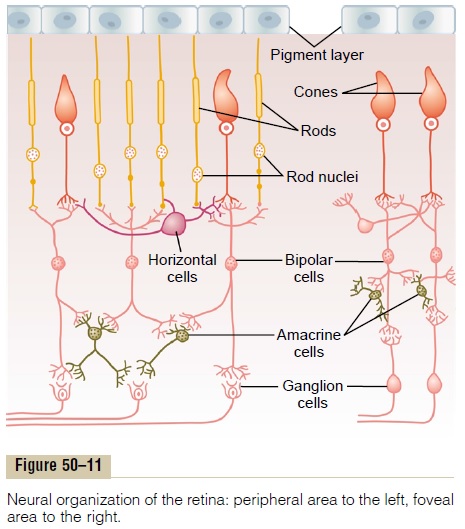
4. The amacrine cells, which transmit signals in two directions, either directly from bipolar cells to ganglion cells or horizontally within the inner plexiform layer from axons of the bipolar cells to dendrites of the ganglion cells or to other amacrine cells
5. The ganglion cells, which transmit output signals from the retina through the optic nerve into the brain
A sixth type of neuronal cell in the retina, not very prominent and not shown in the figure, is the inter-plexiform cell. This cell transmits signals in the retro-grade direction from the inner plexiform layer to the outer plexiform layer. These signals are inhibitory and are believed to control lateral spread of visual signals by the horizontal cells in the outer plexiform layer.
Their role may be to help control the degree of con-trast in the visual image.
The Visual Pathway from the Cones to the Ganglion Cells Func-tions Differently from the Rod Pathway. As is true for manyof our other sensory systems, the retina has both an old type of vision based on rod vision and a new type of vision based on cone vision. The neurons and nerve fibers that conduct the visual signals for cone vision are considerably larger than those that conduct the visual signals for rod vision, and the signals are con-ducted to the brain two to five times as rapidly. Also, the circuitry for the two systems is slightly different, as follows.
To the right in Figure 50–11 is the visual pathway from the foveal portion of the retina, representing the new, fast cone system. This shows three neurons in the direct pathway: (1) cones, (2) bipolar cells, and (3) ganglion cells. In addition, horizontal cells transmit inhibitory signals laterally in the outer plexiform layer, and amacrine cells transmit signals laterally in the inner plexiform layer.
To the left in Figure 50–11 are the neural connec-tions for the peripheral retina, where both rods and cones are present. Three bipolar cells are shown; the middle of these connects only to rods, representing the type of visual system present in many lower animals. The output from the bipolar cell passes only to amacrine cells, which relay the signals to the ganglion cells. Thus, for pure rod vision, there are four neurons in the direct visual pathway: (1) rods, (2) bipolar cells, (3) amacrine cells, and (4) ganglion cells. Also, horizontal and amacrine cells provide lateral connectivity.
The other two bipolar cells shown in the peripheral retinal circuitry of Figure 50–11 connect with both rods and cones; the outputs of these bipolar cells pass both directly to ganglion cells and by way of amacrine cells.
Neurotransmitters Released by Retinal Neurons. Not all theneurotransmitter chemical substances used for synap-tic transmission in the retina have been entirely delin-eated. However, both the rods and the cones release glutamate at their synapses with the bipolar cells.Histological and pharmacological studies have shown there to be many types of amacrine cells secreting at least eight types of transmitter substances, including gamma-aminobutyric acid, glycine, dopamine, acetyl-choline, and indolamine, all of which normally functionas inhibitory transmitters. The transmitters of the bipolar, horizontal, and interplexiform cells are unclear, but at least some of the horizontal cells release inhibitory transmitters.
Transmission of Most Signals Occurs in the Retinal Neurons by Electrotonic Conduction, Not by Action Potentials. The onlyretinal neurons that always transmit visual signals by means of action potentials are the ganglion cells, and they send their signals all the way to the brain through the optic nerve. Occasionally, action potentials have also been recorded in amacrine cells, although the importance of these action potentials is questionable. Otherwise, all the retinal neurons conduct their visual signals by electrotonic conduction, which can be explained as follows.
Electrotonic conduction means direct flow of elec-tric current, not action potentials, in the neuronal cyto-plasm and nerve axons from the point of excitation all the way to the output synapses. Even in the rods and cones, conduction from their outer segments, where the visual signals are generated, to the synaptic bodies is by electrotonic conduction. That is, when hyperpo-larization occurs in response to light in the outer segment of a rod or a cone, almost the same degree of hyperpolarization is conducted by direct electric current flow in the cytoplasm all the way to the synap-tic body, and no action potential is required. Then, when the transmitter from a rod or cone stimulates a bipolar cell or horizontal cell, once again the signal is transmitted from the input to the output by direct elec-tric current flow, not by action potentials.
The importance of electrotonic conduction is that it allows graded conduction of signal strength. Thus, for the rods and cones, the strength of the hyperpolariz-ing output signal is directly related to the intensity of illumination; the signal is not all or none, as would be the case for each action potential.
Lateral Inhibition to Enhance Visual Contrast— Function of the Horizontal Cells The horizontal cells, shown in Figure 50–11, connect laterally between the synaptic bodies of the rods and cones, as well as connecting with the dendrites of the bipolar cells. The outputs of the horizontal cells arealways inhibitory. Therefore, this lateral connectionprovides the same phenomenon of lateral inhibition that is important in all other sensory systems—that is, helping to ensure transmission of visual patterns with proper visual contrast. This phenomenon is demon-strated in Figure 50–12, which shows a minute spot of light focused on the retina. The visual pathway from
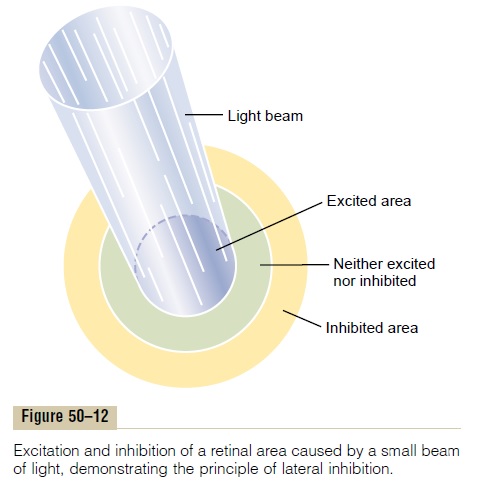
the centralmost area where the light strikes is excited, whereas an area to the side is inhibited. In other words, instead of the excitatory signal spreading widely in the retina because of spreading dendritic and axonal trees in the plexiform layers, transmission through the hor-izontal cells puts a stop to this by providing lateral inhibition in the surrounding areas. This is essential to allow high visual accuracy in transmitting contrast borders in the visual image.
Some of the amacrine cells probably provide addi-tional lateral inhibition and further enhancement of visual contrast in the inner plexiform layer of the retina as well.
Excitation of Some Bipolar Cells and Inhibition of Others—The Depolarizing and Hyperpolarizing Bipolar Cells
Two types of bipolar cells provide opposing excitatory and inhibitory signals in the visual pathway: (1) the depolarizing bipolar cell and (2) the hyperpolarizing bipolar cell. That is, some bipolar cells depolarizewhen the rods and cones are excited, and others hyperpolarize.
There are two possible explanations for this differ-ence. One explanation is that the two bipolar cells are of entirely different types—one responding by depo-larizing in response to the glutamate neurotransmitter released by the rods and cones, and the other respond-ing by hyperpolarizing.The other possibility is that one of the bipolar cells receives direct excitation from the rods and cones, whereas the other receives its signal indirectly through a horizontal cell. Because the hori-zontal cell is an inhibitory cell, this would reverse the polarity of the electrical response.
Regardless of the mechanism for the two types of bipolar responses, the importance of this phenomenon is that it allows half the bipolar cells to transmit positive signals and the other half to transmit negative signals. We shall see later that both positive and nega-tive signals are used in transmitting visual information to the brain.
Another important aspect of this reciprocal relation between depolarizing and hyperpolarizing bipolar cells is that it provides a second mechanism for lateral inhibition, in addition to the horizontal cell mechanism. Because depolarizing and hyperpolarizing bipolar cells lie immediately against each other, this provides a mechanism for separating contrast borders in the visual image, even when the border lies exactly between two adjacent photoreceptors. In contrast, the horizontal cell mechanism for lateral inhibition oper-ates over a much greater distance.
Amacrine Cells and Their Functions
About 30 types of amacrine cells have been identified by morphological or histochemical means. The func-tions of about half a dozen types of amacrine cells have been characterized, and all of them are different. One type of amacrine cell is part of the direct pathway for rod vision—that is, from rod to bipolar cells to amacrine cells to ganglion cells.
Another type of amacrine cell responds strongly at the onset of a continuing visual signal, but the response dies rapidly.
Other amacrine cells respond strongly at the offset of visual signals, but again, the response dies quickly.
Still other amacrine cells respond when a light is turned either on or off, signaling simply a change in illumination, irrespective of direction.
Still another type of amacrine cell responds to movement of a spot across the retina in a specific direction; therefore, these amacrine cells are said to be directional sensitive.
In a sense, then, many or most amacrine cells are interneurons that help analyze visual signals before they ever leave the retina.
Related Topics