Chapter: Microbiology and Immunology: Virology, Virus: General Properties of Viruses
Morphology of Viruses
Morphology of Viruses
The extracellular, infectious viral particle is called the virion:
1. It is a complete, fully developed infectious viral particle composed of nucleic acid surrounded by a protein coat. The latter protects it from the environment and is a vehicle of transmission from one host to another. The viruses are classified on the basis of differences in structure of these coats.
2. The virion may be enveloped by being surrounded by a mem-brane or may be nonenveloped, without being surrounded by a membrane.
3. The virion may also contain essential or accessory enzymes or other proteins.
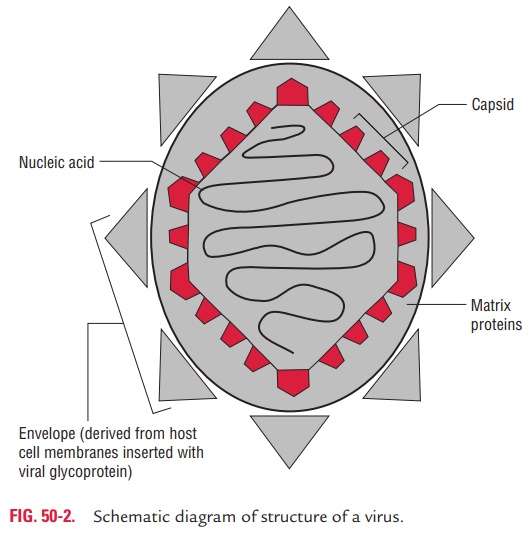
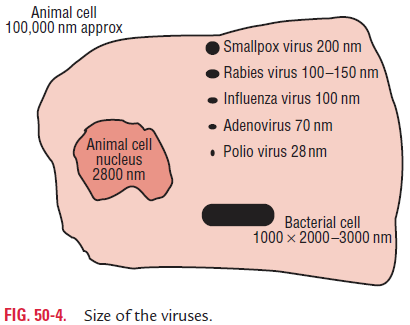
Size
The clinically important viruses vary widely in their size (Fig. 50-1). They range from as small as 20-nm viruses (picor-naviruses) to as large as 300-nm viruses (poxvirus). Passing the viruses through collodion membrane filter with different pore sizes was the earliest method of determining the size of the virus. Subsequently, ultracentrifugation method was used to determine the size of the viruses by calculating from the rate of sedimentation of virus in the ultracentrifuge. Electron microscopy is the most recent method for determining the size as well as the shape of the virus.
Structure and Symmetry of Virus
◗ Viral structure
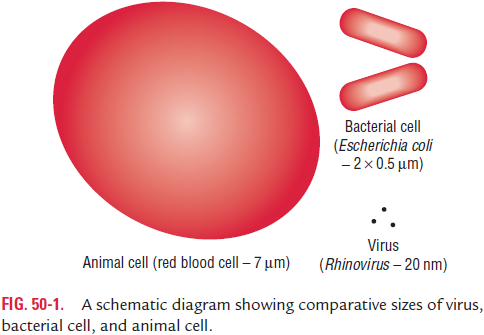
The virion consists of a nucleic acid core, the genome, surrounded by a protein coat, the capsid (Fig. 50-2). The capsid together with the enclosed nucleic acid is known as the nucleocapsid. Some viruses are surrounded by envelopes.
The capsid
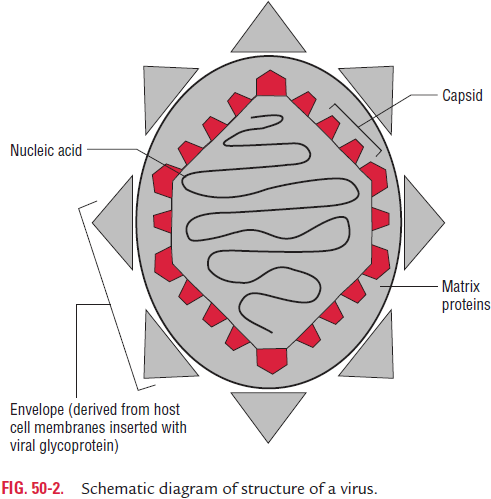
The nucleic acid of a virus is surrounded by a protein coat called the capsid. Each capsid is composed of a large number of protein subunits (polypeptides) called capsomeres, which form its morphological units. The polypeptide molecules compos-ing the capsomeres are of a single type in some viruses, while in other viruses several types may be present. The arrangement of capsomeres is characteristic of a particular type of virus.
The structure of the viral capsid is best demonstrated by X-ray crystallography or electron microscopy. On the basis of capsid structure, the viruses can be classified into different morpho-logical types as follows:
Helical viruses: The helical viruses appear rod-like and may berigid or flexible. The viral genome is found within hollow cylin-drical capsid that has a helical structure. The examples of heli-cal viruses include rabies virus, Ebola hemorrhagic virus, etc.
Polyhedral viruses: The polyhedral viruses appear as many-sided viruses. The viruses consist of capsids in the shape of an icosahedron. It is a regular polyhedron with 20 triangular faces. The capsomere of each face forms an equilateral triangle. Adenovirus is an example of polyhedral virus in the shape of icosahedron.
Enveloped viruses: The helical and polyhedral viruses whencovered by envelope are called as enveloped helical or enveloped polyhedral viruses, respectively. Influenza virus is an example of enveloped helical virus, and herpes simplex virus is an exam-ple of enveloped polyhedral virus.
Complex viruses: Some viruses, such as viruses of bacteria(e.g., bacteriophages), have complicated structures and are called complex viruses.
The envelope
In some viruses, the capsid is covered by an envelope, such viruses are called enveloped viruses. All of the negative-stranded RNA viruses are enveloped. The viruses that lack envelope are called nonenveloped or naked viruses. Properties of the envel-oped and naked viruses are summarized in Tables 50-2 and 50-3, respectively.


The virion envelope usually consists of lipids, proteins, and glycoproteins. It has a membrane structure similar to cellular membrane of the host cell. The viral envelope does not con-tain any cellular proteins, even though viruses are released from the host cell by an extrusion process that coats the virus with a layer of host cell plasma membrane that becomes the viral envelope. In most cases, the envelope contains proteins that are determined and encoded by viral nucleic acid. The lipid compo-nent of the envelope is usually of host cell origin.
Depending on the virus, the envelopes of the viruses may or may not be covered by spikes. The spikes are glycoprotein-like projections on the outer surface of the envelope. Most spikes act as viral attachment protein (VAP).
· The VAP that binds to red blood cells is called hemagglu-tinin. The ability of certain viruses, such as influenza virus to agglutinate red blood cells is due to the presence of these hemagglutinins. The process is called hemagglutination and it forms the basis of hemagglutination inhibition test used in the viral serology.
· The VAPs in some viruses perform different functions, such as neuraminidase activity of influenza virus, fusion glyco-protein of paramyxovirus and C3b receptor associated with herpes simplex virus.
The structural components of envelope remain biologically active only in aqueous solutions and are readily destroyed by drying or on treatment with acids, detergents, and solvents, such as ether, leading to inactivation of virus. They are rapidly killed in stomach due to sensitivity of enveloped components to gastric acidity. Therefore, most of the enveloped viruses are usually transmitted through body fluids, such as blood, respi-ratory droplets, tissue exudates, etc.
◗ Viral symmetry
Three types of symmetry are observed depending on the arrangement of the capsid around the nucleic acid core (genome). These are (a) icosahedral (cubical), (b) helical, and (c) complex symmetry.
Icosahedral symmetry: Two types of capsomeres constitutethe icosahedral capsule. They are the pentagonal capsomeres or the vertices (pentons) and hexagonal capsomeres making up the facets (hexons) (Fig. 50-3). There are always 12 pentons, but the number of hexons varies with the virus group.
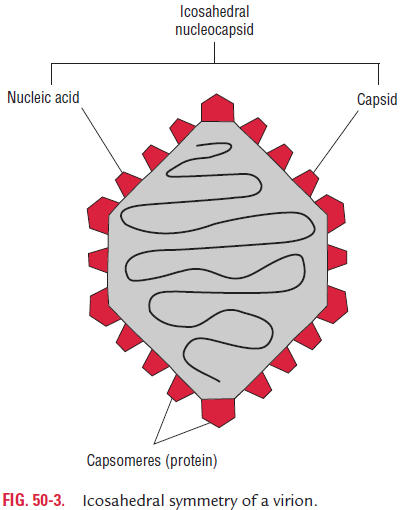
Each penton has fivefold symmetry (pentamer or pentagon) in the shape of an equilateral triangle. This pentamer symmetry is found in simple viruses, such as the picornaviruses and par-voviruses. In picornaviruses, each pentamer is made up of five protomers, each of which is composed of three subunits of four different proteins.
The hexamer symmetry is usually found in large capsid virions, such as herpesviruses and adenoviruses. Hexons are made up of certain structurally distinct capsomeres between the pentons at the vertices. The presence of hexon extends the icosahedral and is called an icosadeltahedral. The adenovirus nucleocapsid has 12 pentons and 240 hexons, whereas the herpesvirus nucleocapsid has 12 pentons and 150 hexons sur-rounded by an envelope.
Helical symmetry: The nucleic acid and the capsomeres arewound together to form a spherical or spiral tube. The viruses with helical structure usually appear as rods and their capso-meres self-assemble on the RNA genome into rods extending to the length of the genome. These capsomeres cover and protect the RNA. The tubular nucleocapsid structure may be rigid as in tobacco mosaic virus, but may be pliable and may be coiled on itself in case of some other animal viruses. Helical nucleo-capsids are usually demonstrated within the envelope of most negative-stranded RNA viruses.
Complex symmetry: Some viruses may not exhibit either ico-sahedral or helical symmetry but instead may exhibit a complex symmetry. For example, poxvirus shows a complex symmetry.
◗ Shape
Most of the enveloped viruses are round or pleomorphic with exception of poxvirus and rhabdovirus. Rhabdovirus is a bullet-shaped virus, whereas poxvirus is brick shaped.
Viral Nucleic Acid, Proteins, and Lipids
◗ Viral nucleic acid
The genome of the virus consists of either DNA or RNA but never both (Fig. 50-4). The DNA can be single stranded or dou-ble stranded. Depending on the virus, the DNA can be linear or circular. The RNA can be either positive sense (1) like mRNA or negative sense (2), double stranded (1/2), or ambiguous (containing 1 and 2 regions of RNA attached to it). In some RNA viruses, such as the influenza virus, the RNA genome is in several separate segments, each segment encoding an individual gene.
The total amount of nucleic acid may vary from a few thousand nucleotides to as many as 250,000 nucleotides.
◗ Viral proteins and lipids
Viruses contain proteins, which constitute capsids. The viral protein protects the nucleic acid as well as determines the antigenic specificity of the virus. In addition, the enveloped viruses contain lipids, which are derived from the host cell membrane.
Susceptibility to Physical and Chemical Agents
◗ Disinfectants
The viruses are usually more resistant than bacteria to chemi-cal disinfectants. Most viruses are relatively resistant to phenol. The oxidizing agents, such as hydrogen peroxide, potassium permanganate, hypochlorite, and organic iodine compounds, are most active antiviral disinfectants. Formaldehyde and b-propiolactone are also active virucidal agents, which arecommonly used for preparation of killed viral vaccines.
The chlorination of drinking water is useful for kill-ing most of the common viruses with exception of hepatitis A and polioviruses. These two viruses are relatively resistant to chlorination.
◗ Temperature
Most of the viruses with few exceptions are highly heat labile. They are inactivated within seconds at 56°C, within minutes at 37°C, and within days at 4°C.
· The viruses such as influenza, measles, and mumps are very labile and may survive outside the host only for a few hours.
· Other viruses, such as polio and hepatitis A, are relatively much stable and may survive for many days, weeks, or even months in the environment.
· Viruses, such as hepatitis B, show resistance to heating at 60°C for 60 minutes; slow viruses, such as scrapie virus, are resistant to autoclaving at 121°C for 15 minutes.
The viruses are stable at low temperature. They can be stored by freezing at 235°C or 270°C. Lyophilization or freeze-drying is useful for long-term storage of the viruses. The poliovirus is an exception, as it does not withstand freeze-drying.
◗ pH
The viruses usually remain viable in a pH range of 5–9, but are sensitive to extremes of acidity and alkalinity. Rhinoviruses are very susceptible to acidic pH, while enteroviruses are highly resistant.
◗ Lipid solvents
Ether, chloroform, and detergents are active against envel-oped viruses but are not active against nonenveloped, naked viruses.
◗ Radiations
The viruses are readily inactivated by sunlight, ultraviolet (UV) radiations, and ionizing radiations.
Related Topics