Chapter: Human Neuroanatomy(Fundamental and Clinical): Internal Structure of Brainstem
Medulla - Internal Structure of Brainstem
The Medulla
Section through the medulla at the level of the pyramidal decussation
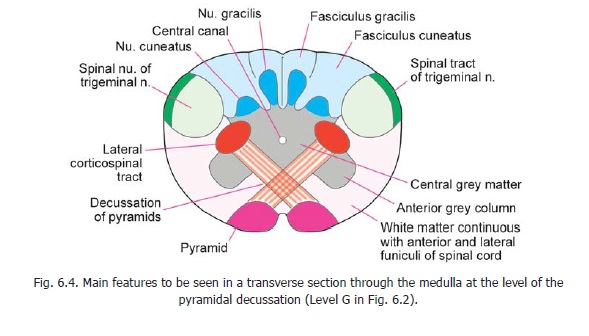
Some features to be seen at this level have been reviewed. The pyramids and their decussation, the nucleus gracilis, the nucleus cuneatus, the spinal nucleus of the trigeminal nerve, the central grey matter, the central canal, and the uppermost part of the ventral grey column have been identified (Fig. 6.4). Now note the following additional features (Fig. 11.1).
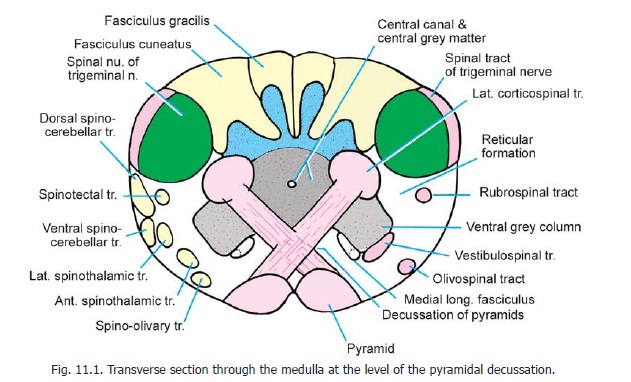
The ventral grey column is separated from the central grey matter by decussating pyramidal fibres. The neurons in it give origin to the uppermost rootlets of the first cervical nerve, and to some fibres in the spinal root of the accessory nerve. The area between the ventral grey column and the spinal nucleus of the trigeminal nerve is occupied by the lower part of the reticular formation.
The main descending fibres to be seen at this level are the corticospinal fibres that form the pyramids. After crossing the middle line these fibres turn downwards in the region lateral to the central grey matter to form the lateral corticospinal tract. We have already seen that those fibres of the pyramids that do not cross descend into the ventral funiculus of the spinal cord to form the ventral corticospinal tract. Other descending tracts to be seen at this level (in the anterolateral part
The tectospinal tract is incorporated within the medial longitudinal fasciculus. Among descending tracts we may also include the spinal tract of the trigeminal nerve, which forms a layer of fibres superficial to the spinal nucleus of this nerve.
The ascending tracts to be seen at this level include the fasciculus gracilis and fasciculus cuneatus which occupy the areas behind the corresponding nuclei; and the spinothalamic, spinocerebellar, spinotectal and spino-olivary tracts that occupy the anterolateral region.
Section through the medulla at the level of the lemniscal decussation
The level represented by this section lies a little above the level of the pyramidal decussation. Some features of a section at this level have already been seen. The central canal surrounded by central grey matter, the medial lemniscus, the pyramids, the nucleus gracilis, the nucleus cuneatus, the spinal nucleus of the trigeminal nerve, and the reticular formation have been identified.
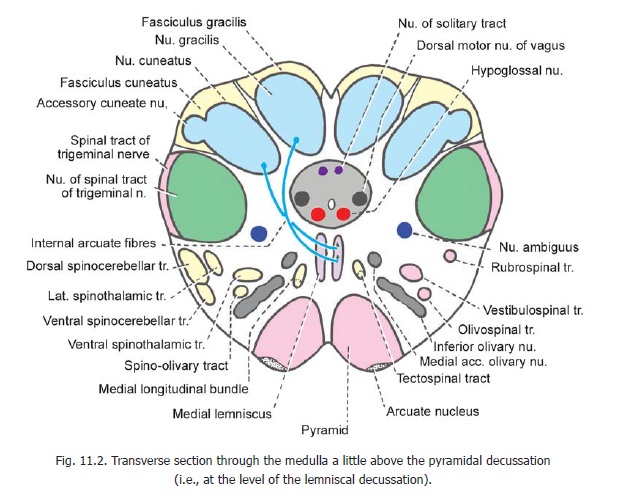
The nucleus gracilis and the nucleus cuneatus are much larger than at lower levels (Fig. 11.2). Internal arcuate fibres arising in these nuclei arch forwards and medially around the central grey matter to cross the middle line. These crossing fibres constitute the lemniscal (or sensory)decussation. Having crossed the middle line these fibres turn cranially to constitute the medial lemniscus. As the fibres from the nucleus gracilis and the nucleus cuneatus pass forwards they crosseach other so that the fibres from the nucleus gracilis come to lie ventral to those from the nucleus cuneatus. The most medial fibres (from the legs) come to lie most anteriorly in the medial lemniscus. These are followed by fibres from the trunk and from the upper limb, in that order. Higher up in the brainstem the medial lemniscus changes its orientation, its long axis (as seen in cross section) becoming transverse (Fig. 11.7). The most anterior fibres become lateral, and the posterior fibres become medial. In its course through the medulla the medial lemniscus is joined by the anterior spinothalamic tract.
It has been shown that fibres in the medial lemniscus are arranged in layers corresponding to spinal segments. Fibres from segment C1 are most medial, and those from S4 are most lateral.
The accessory cuneate nucleus is placed dorsolateral to the cuneate nucleus. It receives proprioceptive impulses from the upper limb through fibres arising in spinal grey matter of cervical segments of the cord. Efferents of the accessory cuneate nucleus constitute the posterior externalarcuate fibres. They reach the cerebellum through the inferior cerebellar peduncle of the same side.
A number of cranial nerve nuclei can be identified at this level. Several of these are present in relation to the central grey matter. The hypoglossal nucleus is located ventral to the central canal just lateral to the middle line. The dorsal vagal nucleus lies dorsolateral to the hypoglossal nucleus. The nucleus of the solitary tract is seen dorsal to the central canal near the middle line. The lower ends of these nuclei of the two sides become continuous with each other to form the commissural nucleus of the vagus. The nucleus ambiguus lies in the reticular formation medial to the spinal nucleus of the trigeminal nerve.
Other masses of grey matter that may be recognised at this level are:
1. the lowest part of the inferior olivary nucleus;
2. the medial accessory olivary nucleus which lies dorsal to the medial part of the inferior olivary nucleus ;
3. the lateral reticular nucleus lying in the lateral part of the reticular formation; and
4. arcuate nuclei lying on the anterior aspect of the pyramids.
The gracile and cuneate fasciculi are much smaller than at lower levels as the fibres of these tracts progressively terminate in the gracile and cuneate nuclei. Other ascending tracts to be seen at this level are the spinothalamic, spinocerebellar, spinotectal and spino-olivary tracts all of which lie in the anterolateral region (Fig. 11.2, left half).
The descending tracts present are (Fig. 11.2, right half) the pyramids, the rubrospinal, vestibulospinal and olivospinal tracts, and the medial longitudinal fasciculus which includes the tectospinal tract.
Section through the medulla at the level of the olive
Some features of a transverse section at this level have been introduced (Fig. 6.6). The floor of the fourth ventricle lined by grey matter, the reticular formation, the spinal nucleus and tract of the trigeminal nerve, the inferior cerebellar peduncle, the inferior olivary nucleus, the medial lemniscus and the pyramids have been briefly considered.
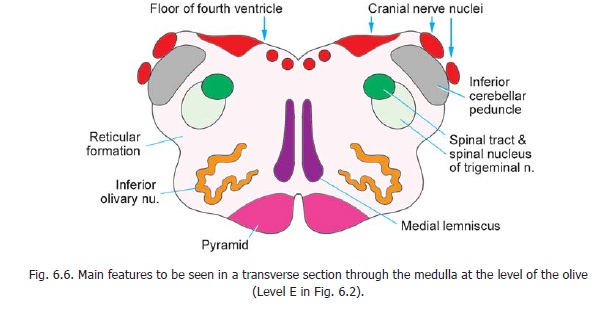
Several cranial nerve nuclei can be recognised in relation to the floor of the fourth ventricle (Fig. 11.3). From medial to lateral side these are the hypoglossal nucleus, the dorsal vagal nucleus, and the vestibular nuclei. The solitary tract and its nucleus lie ventrolateral to the dorsal vagal nucleus. The nucleus ambiguus lies much more ventrally within the reticular formation.
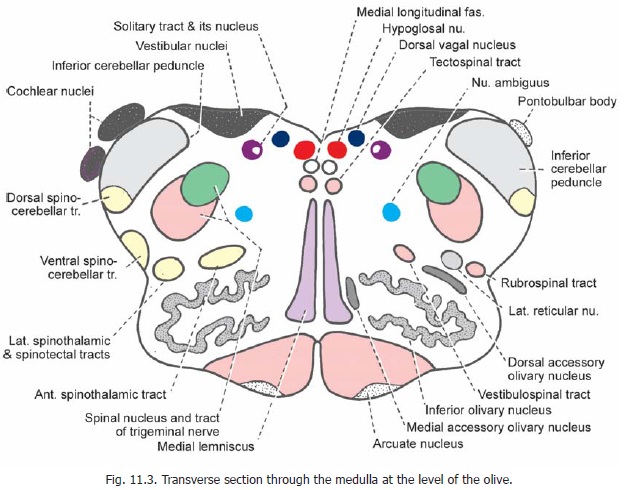
The dorsal and ventral cochlear nuclei can be seen in relation to the inferior cerebellar peduncle. (They are shown schematically in Fig. 11.3. They are actually seen at higher levels of the medulla, near its junction with the pons).
Other masses of grey matter present are the medial and dorsal accessory olivary nuclei (lying medial and dorsal, respectively, to the inferior olivary nucleus), and the lateral reticular nucleus and arcuate nuclei which occupy the same relative positions as at lower levels. The pontobulbar body lies on the dorsolateral aspect of the inferior cerebellar peduncle (Fig. 11.3, right).
The descending tracts to be seen at this level (Fig. 11.3, right half) are the pyramids, the tectospinal, vestibulospinal, and rubrospinal tracts; and the spinal tract of the trigeminal nerve.
The ascending tracts are the medial lemniscus forming an anteroposterior L-shaped band lying next to the middle line; the spinothalamic, spinocerebellar and spino-tectal tracts. At this level the dorsal spino-cerebellar tract lies within the inferior cerebellar peduncle. The ventral spino-cerebellar tract lies more anteriorly near the surface of the medulla. The spinothalamic tracts lie dorsolateral to the inferior olivary nucleus. The medial longitudinal fasciculus lies dorsal to the medial lemniscus.
The connections of the cranial nerve nuclei of the medulla have been described. The reticular formation is considered. Some details about other masses of grey matter to be seen in the medulla are considered below.
Gracile and Cuneate nuclei
We have seen that the gracile and cuneate nuclei receive fibres of the fasciculus gracilis and fasciculus cuneatus respectively; and give origin to the internal arcuate fibres which form the medial lemniscus, ultimately reaching the thalamus. However, the gracile and cuneate nuclei are not to be regarded as simple relay stations. The neurons in them are of various types. In both nuclei an upper reticularregion can be distinguished. Apart from receiving fibres derived from the dorsal nerve roots of spinalnerves, this region receives fibres originating in spinal grey matter. It also receives fibres descending from the sensory cortex: these could presumably inhibit or facilitate conduction through the gracile and cuneate nuclei. Some fibres from these nuclei descend to the spinal grey matter (constituting a feedback circuit).
The arrangement of neurons in the gracile and cuneate nuclei is somatotropic. The upper (reticular) parts of the nuclei receive proprioceptive inputs. The lower parts of the nuclei receive impulses arising (mainly) in skin.
In the cuneate nucleus there is a middle zone called the pars rotunda. Within the pars rotunda distinct areas which receive impulses from individual digits, palm and dorsum of the hand, and from the forearm can be distinguished.
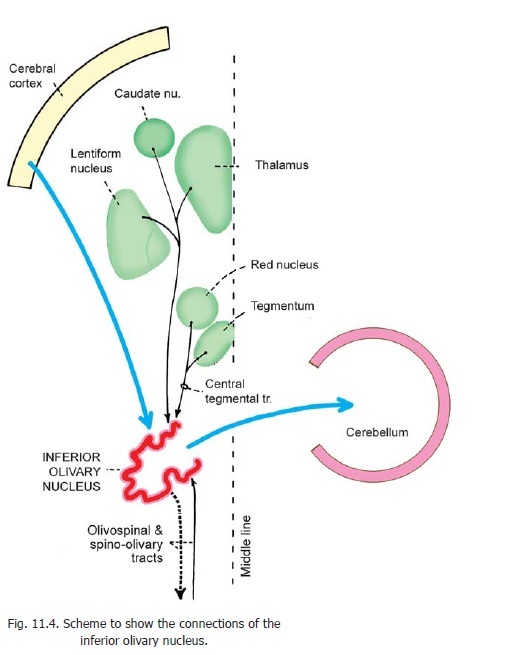
Connections of the Inferior Olivary Complex
The main afferents of the inferior olivary nucleus are from the cerebral cortex and from the spinal cord (Fig. 11.4). The main efferents are to the cerebellar cortex. Details of olivocerebellar connections are considered. An olivospinal tract is traditionally described, but some authorities hold that the inferior olivary nuclei do not send any fibres to the spinal cord. The nucleus may be regarded as a relay station on the cortico-olivo-cerebellar and spino-olivo-cerebellar pathways. Other connections of the nucleus are shown in Fig. 11.4.
The accessory olivary nuclei are connected to the cerebellum by parolivo-cerebellar fibres.
Connections of arcuate nuclei and pontobulbar body
The arcuate nuclei are generally regarded as displaced pontine nuclei. Cortical fibres reach them through the pyramids. These are relayed to the cerebellum by fibres which follow two separate pathways. Some of them wind round the anterior and lateral aspect of the medulla as anterior external arcuate fibres to reach the inferior cerebellar peduncle of the opposite side. Other fibres pass dorsally through the substance of the medulla to reach the floor of the fourth ventricle. Here they run under the ependyma to the inferior cerebellar peduncle of the opposite side as fibres of the striae medullares. Fibres from the arcuate nucleiprobably end in the flocculus of the cerebellum.
Students must distinguish carefully between the stria medullares described above and the stria medullaris thalami present in relation to the wall of the third ventricle.
Like the arcuate nuclei, the pontobulbar body is made up of neurons that represent displaced pontine nuclei. Fibres arising in this body form the circumolivary bundle. These fibres join those from the arcuate nuclei to reach the inferior cerebellar peduncle of the opposite side. Some of them possibly pass through the striae medullares.
Related Topics