Chapter: Essential Clinical Immunology: Basic Components of the Immune System
Major Histocompatibility Complex
MAJOR HISTOCOMPATIBILITY COMPLEX
Human histocompatibility antigens are also known as
human leucocyte antigens (HLA), a term that is synonymous with the MHC complex.
These antigens are cell-surface glycoproteins classified as type I or type II.
They can produce genetic poly-morphism with multiple alleles at each site, thus
permitting a great deal of genetic variability between given individuals (see
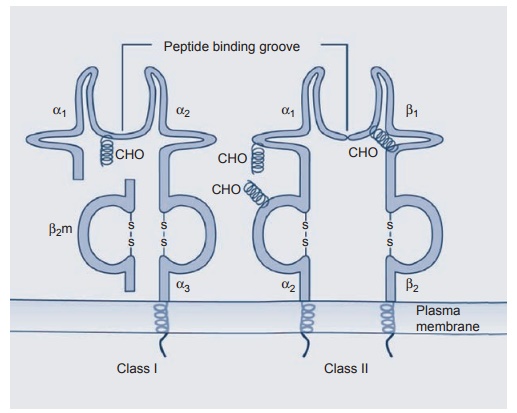
Figure 1.6 Diagrammatic
representation of class I and II MHC antigens with B2 microglobulins
and CHO carbohydrate side chains.
Figure 1.6). This extensive polymorphism is
important when viewed in the context of an immune system that needs to cope
with an ever-increasing range of pathogens. These pathogens in turn are
extremely adept at evading the immune system. Thus, the battle between invading
microbe and immune recognition is constant and ever changing. Recognition of
antigen by T cells is MHC restricted. Therefore, any given individual is only
able to recognize antigen as part of a complex of antigenic peptide and self.
The importance of this concept is under-scored by
the experiments of Dougherty and Zinkernagel. Using virus-specific (virus 1)
cytotoxic T cells, Figure 1.7 illus-trates their remarkable discovery. If
anti-gen-presenting cells (APCs) of mouse A are mixed with T cells of mouse A
in the context of virus 1 peptides, the T cell responds and kills the virus. If
the MHC complex is from mouse B and the T cells
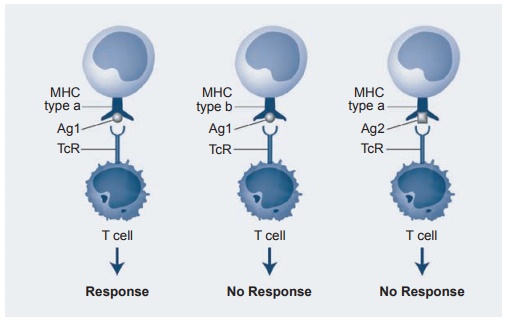
Figure 1.7 MHC restriction of antigen recognition by T cells. If APC and T cell are of the same genetic lineage as virus I, the T cell responds and kills the virus. If APC and T cell are of different lineage, no response occurs. If APC and T cell are of same lineage but virus 2 is present, no response occurs.
The MHC
class I antigens are divided into three groups (A, B, and C), and each group
belongs on a different gene locus on chromosome 6. The products of all three
loci are similar and are made up of a heavy chain (45 kDa) and associated β2
micro-globulin molecule (12 kDa) gene, which resides on chromosome 12. The MHC
class I antigen differences are due to variations in the α chains, the β2
microglobulin being constant. X-ray crystallography studies have shown that as
few as nine amino acids can be tightly bound in the α chain groove.
MHC class
II antigens also exhibit a similar structure with the groove being formed by
the α1 and β1 chains.
Unlike class I antigens present on most nucleated cells, the class II antigens
are restricted
to a few types: macrophages, B cells, and acti-vated T cells. In humans, there
are three groups of class II antigens: namely, HLA-DP, HLA-DQ, and HLA-DR.
Depending
on the nature of the anti-gen (endogenous or exogenous), the MHC response is
different. For example, endog-enous antigens (including viral antigens) are
presented by MHC class I antigen cells exclusively to CD8 cells. The endogenous
antigen is first broken down into small pep-tides and transported by shuttle
proteins called Tap I and Tap II to the endoplasmic reticulum. There they
complex with MHC class I molecules and are delivered to the cell surface for
further processing to the CD8 cells.
In
contrast, MHC class II molecules are held in the endoplasmic reticulum and are
protected from binding to peptides in the lumen (not human) by a protein called
MHC class II associated invariant chain.
Finally,
there are class III antigens, such as complement components C4 and C2,
plus certain inflammatory proteins, such as tumor necrosis factor (TNF), which
are encoded in adjacent areas.
Related Topics