Chapter: Biochemistry: The Citric Acid Cycle
How Pyruvate Is Converted to Acetyl-CoA
How Pyruvate Is Converted to
Acetyl-CoA
Pyruvate can come from several sources, including glycolysis, as we
have seen. It moves from the cytosol into the mitochondrion via a specific
transporter. There, an enzyme system called the pyruvate dehydrogenase complex is responsible for the conversion of
pyruvate to carbon dioxide and the acetyl portion of acetyl-CoA. There is an
ŌĆöSH group at one end of the CoA molecule, which is the point at which the acetyl group is attached. As a
result, CoA is frequently shown in equations as CoA-SH. Because CoA is a thiol
(the sulfur [thio] analog of an alcohol), acetyl-CoA is a thioester, with a sulfur atom replacing an oxygen of the usual
carboxylic ester. This difference is important, since thioesters are
high-energy compounds. In other words, the hydrolysis of thioesters releases
enough energy to drive other reactions. An oxidation reaction precedes the
transfer of the acetyl group to the CoA. The whole process involves several
enzymes, all of which are part of the pyruvate dehydrogenase complex. The
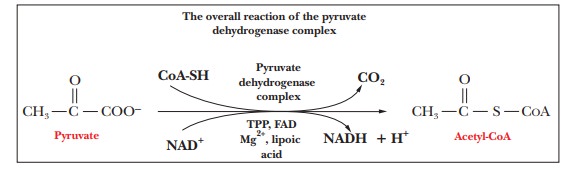
overall reaction
Pyruvate
+ CoA-SH + NAD+ - > Acetyl-CoA + CO2 + H+ +
NADH
is exergonic ( ŌłåG┬░' = ŌĆō33.4 kJ molŌĆō1 = ŌĆō8.0 kcal molŌĆō1), and NADH can then be used to generate ATP via the electron transport chain.
How many enzymes are needed to convert pyruvate to acetyl-CoA?
Five
enzymes make up the pyruvate dehydrogenase complex in mammals. They are pyruvate dehydrogenase (PDH), dihydrolipoyl
transacetylase, dihydrolipoyldehydrogenase, pyruvate dehydrogenase kinase, and pyruvate dehydrogenase phosphatase. The
first three are involved in the conversion of pyruvate to acetyl-CoA. The
kinase and the phosphatase are enzymes used in the control of PDH and are present on a single polypeptide. The
reaction takes place in five steps. Two enzymes catalyze reactions of lipoic acid, a compound that has a
disulfide group in its oxidized form and two sulfhydryl groups in its reduced
form.
Lipoic
acid differs in one respect from other coenzymes. It is a vitamin, rather than
a metabolite of a vitamin, as is the case with many other coenzymes (Table
7.3). (The classification of lipoic acid as a vitamin is open to question.
There is no evidence of a requirement for it in the human diet, but it is
required for the growth of some bacteria and protists.) Lipoic acid can act as
an oxidizing agent; the reaction involves hydrogen transfer, which frequently
accompanies biological oxidationŌĆōreduction reactions. Another reaction of
lipoic acid is the formation of a thioester linkage with the acetyl group
before it is transferred to the acetyl-CoA. Lipoic acid can act simply as an
oxidizing agent, or it can simultaneously take part in two reactionsŌĆöa redox
reaction and the shift of an acetyl group by transesterification.
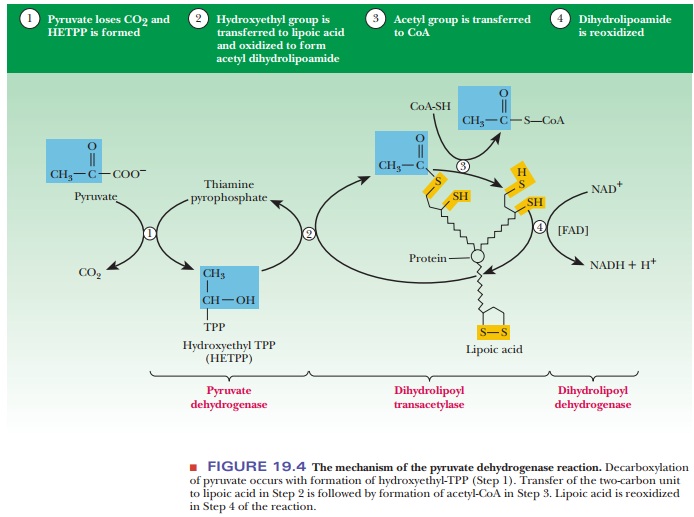
The
first step in the reaction sequence that converts pyruvate to carbon diox-ide
and acetyl-CoA is catalyzed by pyruvate dehydrogenase, as shown in Figure 19.4.
This enzyme requires thiamine pyrophosphate (TPP; a metabolite of vita-min B1,
or thiamine) as a coenzyme. The coenzyme is not covalently bonded to the
enzyme; they are held together by noncovalent interactions. Mg2+ is
also required. We saw the action of TPP as a coenzyme in the conversion of
pyruvate to acetaldehyde, catalyzed by pyruvate decarboxylase. In the pyruvate
dehydrogenase reaction, an ╬▒-keto acid, pyruvate, loses carbon dioxide; the
remaining two-carbon unit becomes covalently bonded to TPP.
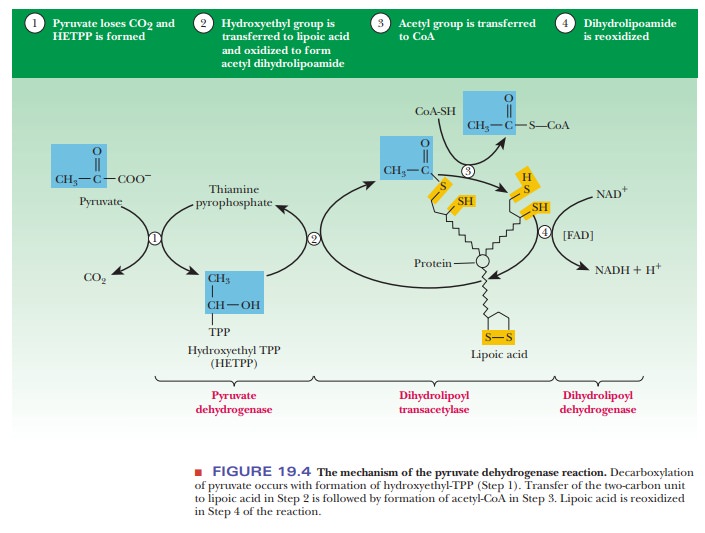
The second step of the reaction is catalyzed by dihydrolipoyl transacetylase. This enzyme requires lipoic acid as a coenzyme. The lipoic acid is covalently bonded to the enzyme by an amide bond to the e-amino group of a lysine side chain. The two-carbon unit that originally came from pyruvate is transferred from the thiamine pyrophosphate to the lipoic acid, and, in the process, a hydroxyl group is oxidized to produce an acetyl group. The disulfide group of the lipoic acid is the oxidizing agent, which is itself reduced, and the product of the reaction is a thioester. In other words, the acetyl group is now covalently bonded to the lipoic acid by a thioester linkage (see Figure 19.4).
The
third step of the reaction is also catalyzed by dihydrolipoyl transacety-lase.
A molecule of CoA-SH attacks the thioester linkage, and the acetyl group is
transferred to it. The acetyl group remains bound in a thioester linkage; this
time it appears as acetyl-CoA rather than esterified to lipoic acid. The
reduced form of lipoic acid remains covalently bound to dihydrolipoyl
transacetylase (see Figure 19.4). The reaction of pyruvate and CoA-SH has now
reached the stage of the products, carbon dioxide and acetyl-CoA, but the
lipoic acid coen-zyme is in a reduced form. The rest of the steps regenerate
the lipoic acid, so further reactions can be catalyzed by the transacetylase.
In the
fourth step of the overall reaction, the enzyme dihydrolipoyl dehydro-genase
reoxidizes the reduced lipoic acid from the sulfhydryl to the disulfide form.
The lipoic acid still remains covalently bonded to the transacetylase enzyme.
The dehydrogenase also has a coenzyme, FAD, that is bound to the enzyme by
noncovalent interactions. As a result, FAD is reduced to FADH2. FADH2
is reoxidized in turn. The oxidizing agent is NAD+, and NADH is the
product along with reoxidized FAD. Enzymes such as pyruvate dehydrogenase are
called flavoproteins because of their attached FADs.
The
reduction of NAD+ to NADH accompanies the oxidation of pyruvate to
the acetyl group, and the overall equation shows that there has been a transfer
of two electrons from pyruvate to NAD+ (Equation 19.1). The
electrons gained by NAD+ in generating NADH in this step are passed
to the electron transport chain (the next step in aerobic metabolism). We shall
see that the transfer of electrons from NADH ultimately to oxygen will give
rise to 2.5 ATP. Two molecules of pyruvate are produced for each molecule of
glucose, so that there will eventually be five ATP from each glucose from this
step alone.
The reaction leading from pyruvate to acetyl-CoA is a complex one that requires three enzymes, each of which has its own coenzyme in addition to NAD+. The spatial orientation of the individual enzyme molecules with respect to one another is itself complex. In the enzyme isolated from E. coli, the arrangement is quite compact, so that the various steps of the reaction can be thoroughly coordinated. There is a core of 24 dihydrolipoyl transacetylase molecules. The 24 polypeptide chains are arranged in eight trimers, with each trimer occupying the corner of a cube. There are 12 ╬▒╬▓ dimers of pyruvate dehydrogenase, and they occupy the edges of the cube. Finally, six dimers of dihydrolipoyl dehydrogenase lie on the six faces of the cube (Figure 19.5). Note that many levels of structure combine to produce a suitable environment for the conversion of pyruvate to acetyl-CoA. Each enzyme molecule in this array has its own tertiary structure, and the array itself has the cubical structure we have just seen.
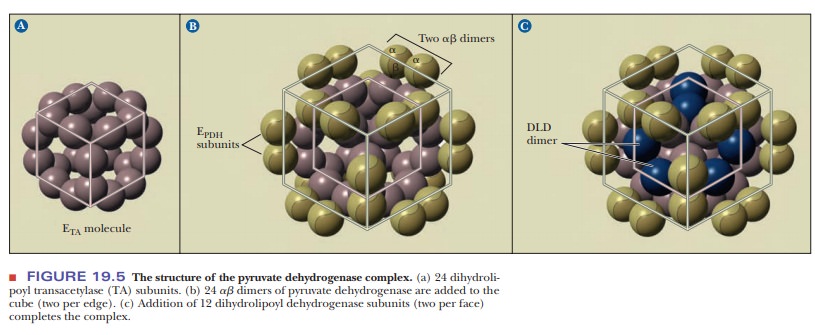
A
compact arrangement, such as the one in the pyruvate dehydrogenase multi-enzyme
complex, has two great advantages over an arrangement in which the vari-ous
components are more widely dispersed. First, the various stages of the reaction
can take place more efficiently because the reactants and the enzymes are so
close to each other. The role of lipoic acid is particularly important here.
Recall that the lipoic acid is covalently attached to the transacetylase enzyme
that occupies a central position in the complex. The lipoic acid and the lysine
side chain to which it is bonded are long enough to act as a ŌĆ£swinging arm,ŌĆØ
which can move to the site of each of the steps of the reaction (Figure 19.4).
As a result of the swinging-arm action, the lipoic acid can move to the
pyruvate dehydrogenase site to accept the two-carbon unit and then transfer it
to the active site of the transacetylase. The acetyl group can then be
transesterified to CoA-SH from the lipoic acid. Finally, the lipoic acid can
swing to the active site of the dehydrogenase so that the sulfhydryl groups can
be reoxidized to a disulfide.
A second
advantage of a multienzyme complex is that regulatory controls can be applied
more efficiently in such a system than in a single enzyme mol-ecule. In the
case of the pyruvate dehydrogenase complex, controlling factors are intimately
associated with the multienzyme complex itself.
Summary
The
two-carbon unit needed at the start of the citric acid cycle is obtained by
converting pyruvate to acetyl-CoA.
This conversion requires the three primary enzymes of the pyruvate dehydro-genase complex, as well as the cofactors TPP, FAD, NAD+, and lipoic acid.
The
overall reaction of pyruvate dehydrogenase is the conversion of pyru-vate, NAD+,
and CoA-SH to acetyl-CoA, NADH + H+, and CO2.
Related Topics