Chapter: Genetics and Molecular Biology: Induction, Repression, and the araBAD Operon
Genetics of the Arabinose System
Genetics of the Arabinose System
Most mutations in the arabinose operon
generate the expected pheno - types. The araB,
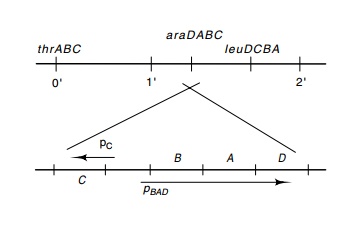
araA, and araD genes lie in one transcriptional unit served by promoter pBAD (Fig. 12.4), and
mutations in these genes abolish the activity of the enzyme in question as well
as leave the cells arabinose-negative. Mutations in araD, however, have the useful prop-erty that they make the cells
sensitive to the presence of arabinose due to their resulting accumulation of
ribulose phosphate. This type of sensitivity is not an isolated case, for high
levels of many sugar phos-phates in many types of cells are toxic or growth
inhibitory.
The
arabinose-sensitivity of AraD- cells provides a simple way to
isolate mutations in the arabinose genes. AraD- mutants that have
further mutated to become resistant to arabinose contain mutations preventing
the accumulation of ribulose phosphate. Each cell capable of growing into a
colony must contain a secondary mutation in the
Figure
12.4 Fine structure of theE.
coli genome in the region of the
araBAD genes, the ara promoters,and
the directions of transcription of the genes. In the top, the genes are drawn
in the conventional ori-entation of the E.
coli genetic map, and in the bottom,
the arabinose genes have been inverted to con-form with the conventional usage,
in which the major transcription is rightward.
arabinose
operon in addition to the AraD- mutation. Such mutants are isolated
by spreading large numbers of AraD- cells onto plates contain-ing
arabinose and some other source of carbon and energy, such as glycerol or yeast
extract. The few cells that grow into colonies contain additional mutations in araC, B, or A. This scheme does not yield transport-negative mutants since both
transport systems would have to be inactivated before the cells would be
arabinose-resistant, and such double mutants are too rare to be detected.
The
behavior of mutations in one of the arabinose genes was para-doxical. The cells
became arabinose-negative, but no enzymatic activity could be associated with
the gene. In fact, none seemed necessary because the other arabinose gene
products performed all the required metabolic conversions. Additionally, cells
with mutations in this gene, araC,
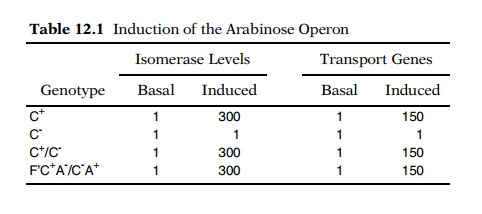
had the strange property of not possessing any of the arabinose-induced enzymes
or active transport systems (Table 12.1). If the gene product were not any of
the logical proteins, perhaps it wasn’t a protein. This wasn’t possible,
however, because nonsense mutations existed in the araC gene. Only genes encoding proteins can possess nonsense
mutations. Therefore, the araC gene
product had to be a protein.
Formally,
the behavior of the araC mutants was
consistent with several regulatory mechanisms. First, contrary to expectations
preva-lent at the time of this work, araC
could code for a positive regulator,
Figure
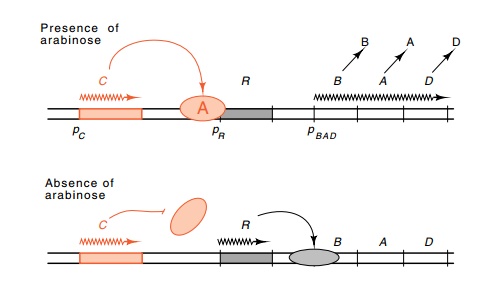
12.5 A double-negative regulation
system in which C looks like anactivator in genetic and physiological tests. C
is a repressor of R synthesis, R is a repressor of the BAD operon.
one whose presence is necessary for synthesis of
the other ara gene products. Second, araC could code for a component directly
involved with the uptake of arabinose into cells. Its absence would mean that
the intracellular level of arabinose would never become high enough to permit
derepression of the arabinose genes. Third, araC
could be part of a double-negative regulation system (Fig. 12.5). That is, AraC
protein in the presence of arabinose could repress the synthesis of the true
repressor of the arabinose operon. In this case, if araC were inactive or arabinose were absent, then the repressor
would be synthesized and the arabinose enzymes would not be synthesized.
Related Topics