Chapter: Plant Biology : Interactions between plants and other organisms
Fungal pathogens and endophytes
FUNGAL PATHOGENS AND ENDOPHYTES
Key Notes
Fungal diseases
Fungi are important saprophytes that recycle dead plant material. They also cause a wide range of plant disease. Fungal spores germinate on a leaf surface or wound site and form a mycelium through the plant. Their life cycles may involve more than one host plant (e.g. wheat rust, which is heteroecious) or a single host plant (e.g. smut fungi, which are autoecious).
Plant defenses against fungal diseases
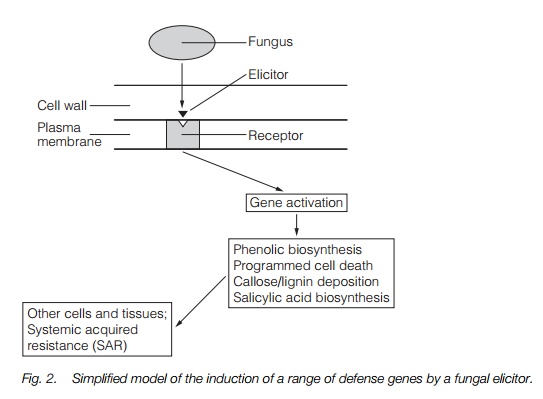
Defenses may be mechanical, preventing spore adherence or penetration, or chemical, e.g. saponins that disrupt fungal membranes. Other defenses are induced when a pathogen attacks, genes being activated by an elicitor binding to a plasma membrane receptor. In a hypersensitive response, cells fill with phenolic compounds and form a necrotic lesion. A lignin or callose barrier may form. In many species, antimicrobial phytoalexins are produced. Systemic acquired resistance occurs when resistance is enhanced as a result of an infection. Fungi may stimulate changes in metabolism or phytohormone levels in the plant leading to changes in growth form.
Fungal endophytes
Fungi are often found growing freely within plants which have no, or minimal symptoms. They can be particularly dangerous to humans as they can be very toxic. Best known is ergot, a fungus growing in rye that generates a toxic precursor to LSD. Some may become parasites as the plant ages.
Fungal diseases
Different fungal species possess a wide range of methods capable of degrading a vast range of materials, including lignin and cellulose. Many fungi are saprophytes and live by degrading material from plants that have already died, and these are essential in soil formation. Others are pathogens that cause a wide range of plant diseases resulting in significant losses in agriculture. Table 1 gives some examples of fungal diseases and the organisms that cause them.

Infection occurs when a spore germinates on a leaf surface or wound site. The fungus penetrates, often through stomata, and forms fine threads of cells (hyphae) which ramify to form a mat (mycelium) throughout the plant. Infection in cells frequently resembles the arbuscules of endomycorrhizae .
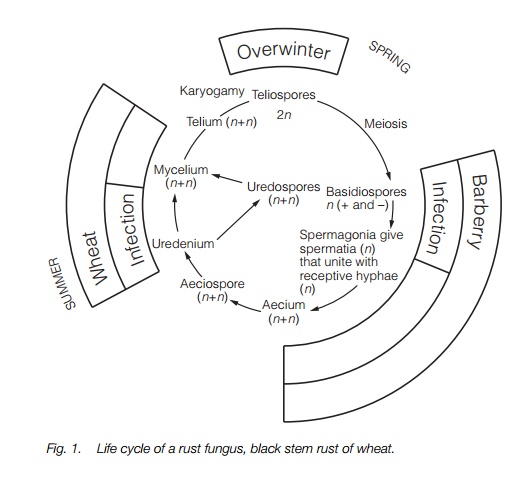
Fungi have complex life cycles, which may involve one or more host plant (heteroecious life cycle). Wheat rust, Puccinia graminis for instance, aBasidiomycete, invades two host plants. Commonly, the first host is the barberry (Berberis sp.) from which aeciospores (containing two nuclei, one + strain, the other – strain) are produced which infect the wheat. The wheat then produces uredospores, which spread the infection in the wheat field. The fungus overwinters as diploid teliospores, produced when the two (+ and –) nuclei fuse (karyogamy). Meiosis occurs in spring to releasebasidiospores (+ or – strain) which then infect the barberry host (Fig. 1). Smut fungi have a simpler life cycle involving only one host (autoecious) and produce masses of teliospores (the resting stage of the fungus). They are known to infect at least 4000 species of flowering plants, many of economic importance.
Plant defenses against fungal diseases
Some defenses are mechanical barriers, for instance the waxy cuticle, which prevents spores from adhering to the leaf or the fungus from penetrating the leaf. Pathogenic fungi can often only infect a wound site or by entering the leaf through stomata. Other defenses are endogenous chemicals, for instance the saponins are triterpenes produced by plants which disrupt the membranes of fungi. Other defenses against fungi are not induced until a pathogen attacks. This involves expression of defense genes which are activated by a signaling cascade within the plant. This is triggered by the binding of a product of the pathogen (an elicitor) to a receptor in the plant plasma membrane (Fig. 2). In some instances, a hypersensitive response occurs, where cells around the fungus fill with phenolic compounds and die to form a necrotic lesion. This isolates the point of infection and may stop further spread of the fungus. The infection site may also be surrounded by deposits of lignin or callose synthesized by surrounding cells which form a physical barrier to the spread of the infection. Another group of antimicrobial compounds, the phytoalexins are produced in many species as a response to infection. Phytoalexin is a generic name for a range of compounds that differ between species. After infection, expression of genes for the enzymes of their biosynthetic pathway is initiated and high levels of phytoalexins accumulate. An infection which has been successfully overcome may lead to the plant being more resistant to further infection. This systemic acquired resistance involves the spread of signaling molecules (possibly salicylic acid) within the plant (Fig. 2).
Some fungi stimulate changes in the metabolism of the plant. The plant’s respiration rate increases and sometimes it shifts more to the pentose phosphate pathway rather than the tricarboxylic acid pathway . This allows for the production of more nucleic acid (which use pentoses) and NADPH, important for the fungus. The fungus may prevent much or all of the export of carbohydrate from leaves to storage organs. The fungus may also stimulate changes in phytohormone levels which lead to unusual growths, one of the most common being ‘witches brooms’, a dense group of short stems in many trees such as birches (Betula spp.).
Fungal Endophytes
Plants are often found with fungal hyphae growing freely within their tissues. They often produce either no, or minimal symptoms of their presence and may provide advantages to the host in deterring herbivores and other pathogens.
These fungal endophytes can be particularly dangerous to humans. One of the best known, ergot (Claviceps purpurea), grows in rye and generateslysergic acidamide, the precursor of lysergic acid diethylamine (LSD). LSD is hallucinogenic and fatal in relatively small doses and throughout history, communities have been devastated by outbreaks undetected in the rye harvest. Other endophytes include Sphacelia typhina, an ascomycete which grows in the prairie grass Festucaarundinacea and causes cattle disease and death. Detection and elimination of endophytes in agriculture and food processing is an important area.
There is no absolute distinction between fungal endophytes and parasites and some fungi may remain as endophytes for many years before starting to damage a plant significantly. Many only reproduce sexually once the plant or the infected part is killed.
Related Topics