Chapter: Basic & Clinical Pharmacology : Introduction to the Pharmacology of Central Nervous System (CNS) Drugs
Cellular Organization of the Brain
CELLULAR ORGANIZATION OF THE
BRAIN
Most
of the neuronal systems in the CNS can be divided into two broad categories: hierarchical systems and nonspecific or diffuse neuronal systems.
Hierarchical Systems
Hierarchical
systems include all the pathways directly involved in sensory perception and
motor control. The pathways are generally clearly delineated, being composed of
large myelinated fibers that can often conduct action potentials at a rate of
more than 50 m/s. The information is typically phasic and occurs in bursts of
action potentials. In sensory systems, the information is processed
sequentially by successive integrations at each relay nucleus on its way to the
cortex. A lesion at any link incapacitates the system. Within each nucleus and
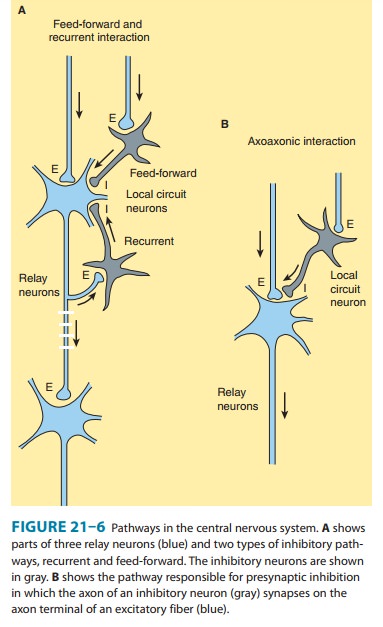
in the cortex, there are two types of cells: relay or projection neurons and local circuit neurons (Figure 21–6A).
The projection neurons that form the intercon-necting pathways transmit signals
over long distances. The cell bodies are relatively large, and their axons emit
collaterals that arborize extensively in the vicinity of the neuron. These
neurons are excitatory, and their synaptic influences, which involve
iono-tropic receptors, are very short-lived.
The
excitatory transmitter released from these cells is, in most instances, glutamate. Local circuit neurons are
typically smaller than projection neurons, and their axons arborize in the
immedi-ate vicinity of the cell body. Most of these neurons are inhibitory, and
they release either GABA or glycine. They synapse primarily on the
cell body of the projection neurons but can also synapse on the dendrites of
projection neurons as well as with each other. Two common types of pathways for
these neurons (Figure 21–6A) include recurrent feedback pathways and feed-forward
pathways. A special class of local circuit neurons in the spinal cord forms
axoaxonic synapses on the terminals of sensory axons (Figure 21–6B). In some
sensory pathways such as the retina and olfactory bulb, local circuit neurons
may actually lack an axon and release neurotransmitter from dendritic synapses
in a graded fashion in the absence of action potentials.
Although
there is a great variety of synaptic connections in these hierarchical systems,
the fact that a limited number of trans-mitters are used by these neurons
indicates that any major phar-macologic manipulation of this system will have a
profound effect on the overall excitability of the CNS. For instance,
selectively blocking GABAA receptors with a drug such as picrotoxin
results in generalized convulsions. Thus, although the mechanism of action of
picrotoxin is specific in blocking the effects of GABA, the overall functional
effect appears to be quite nonspecific, because GABA-mediated synaptic
inhibition is so widely utilized in the brain.
Nonspecific or Diffuse Neuronal
Systems
Neuronal
systems that contain one of the monoamines— norepinephrine, dopamine, or
5-hydroxytryptamine (serotonin)— provide examples in this category. Certain
other pathways emanating from the reticular formation and possibly some
peptide-containing pathways also fall into this category. These systems differ
in fundamental ways from the hierarchical systems, and the noradrenergic
systems serve to illustrate the differences.
Noradrenergic
cell bodies are found primarily in a compact cell group called the locus
caeruleus located in the caudal pon-tine central gray matter. The number of
neurons in this cell group is small, approximately 1500 on each side of the
brain in the rat.
Because
these axons are fine and unmyelinated, they conduct very slowly, at about 0.5
m/s. The axons branch repeatedly and are extraordinarily divergent. Branches
from the same neuron can innervate several functionally different parts of the
CNS. In the neocortex, these fibers have a tangential organization and
there-fore can monosynaptically influence large areas of cortex. The pattern of
innervation by noradrenergic fibers in the cortex and nuclei of the
hierarchical systems is diffuse, and these fibers form a very small percentage
of the total number in the area. In addi-tion, the axons are studded with
periodic enlargements called varicosities, which contain large numbers of
vesicles. In some instances, these varicosities do not form synaptic contacts,
sug-gesting that norepinephrine may be released in a rather diffuse manner, as
occurs with the noradrenergic autonomic innervation of smooth muscle. This
indicates that the cellular targets of these systems are determined largely by
the location of the receptors rather than by the location of the release sites.
Finally, most neu-rotransmitters utilized by diffuse neuronal systems,
including norepinephrine, act—perhaps exclusively—on metabotropic receptors and
therefore initiate long-lasting synaptic effects. Based on these observations,
it is clear that the monoamine sys-tems cannot be conveying topographically
specific types of infor-mation; rather, vast areas of the CNS must be affected
simultaneously and in a rather uniform way. It is not surprising, then, that
these systems have been implicated in such global func-tions as sleeping and
waking, attention, appetite, and emotional states.
Related Topics