Chapter: Plant Biology : Metabolism
C3 and C4 plants and CAM
C3 AND C4 PLANTS AND CAM
Key Notes
CO2 concentration and water conservation
Land plants have to open their stomata to admit CO2. This means water loss is inevitable while photosynthesis occurs. Atmospheric concentrations of CO2 do not saturate Rubisco and it therefore functions at less than its maximal rate. C4-photosynthesis and crassulacean acid metabolism (CAM) are two adaptations which decrease water loss.
C4 anatomy
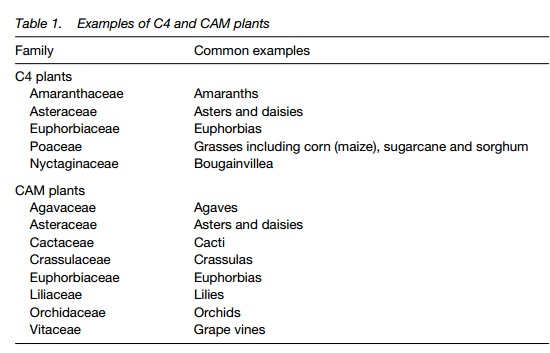
C4 plants show Krantz anatomy, a ring of bundle-sheath cells containing chloroplasts surrounding the leaf veins. Surrounding mesophyll cells also contain chloroplasts and are in close contact with the bundle sheath cells by plasmodesmata.
C4 biochemistry
C4 plants carry out the functions of the Calvin cycle in the bundle sheath cells. Rubisco in these cells is supplied with CO2 released from the C4 compounds malic acid or aspartic acid generated in the mesophyll cells and transported through the plasmodesmata. CO2 is first fixed by phosphoenolpyruvate (PEP)-carboxylase, which has a high affinity for HCO3–. The effect of the system is to increase the efficiency of CO2 fixation and reduce the requirement for stomatal opening.
CAM anatomy
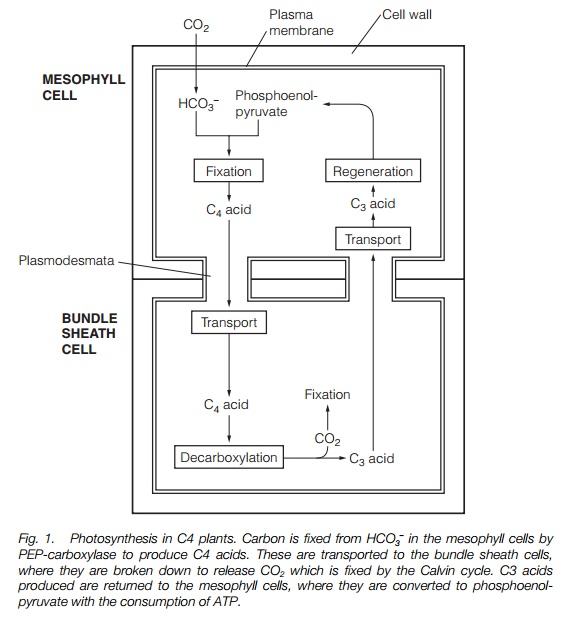
CAM plants show adaptations to drought and are succulents, having fleshy leaves with a minimum surface area to volume ratio. The photosynthetic cells have a substantial vacuole in which C4 acids are stored.
CAM biochemistry
In CAM plants, the stages of CO2 fixation by PEP-carboxylase and of the Calvin cycle occur in the same cells but at different times. CO2 is fixed at night and the C4 compounds formed are stored in the leaf cell vacuole. The C4 compounds break down to release CO2 during the day permitting the Calvin cycle to function when stomata are closed.
Distribution of CAM and C4 photosynthesis
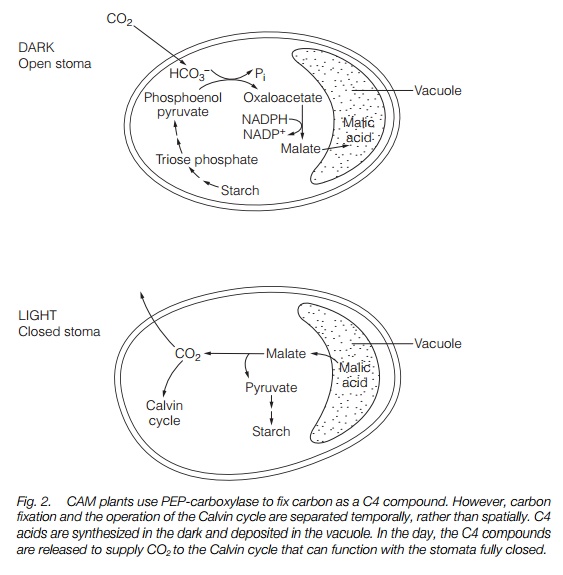
Both C4 and CAM are adaptations to drought occurring in a wide range of species and families, generally from arid zones. Some families of plants contain C3, C4 and CAM members.
CO2 concentration and water conservation
Land plants have a dilemma: allowing a free diffusion of CO2 into leaf cells without excessive water loss. The plant’s environment is often most desiccating when light intensities are highest, meaning that stomata close during times
when CO2 could be used most effectively. In addition, the photosyntheticapparatus of many plants showing C3 photosynthesis, the type of photosynthesis described (so called because its first stage involves formation of two C3 sugars) operates maximally at CO2 concentrations above that found in the atmosphere. Two adaptations have been described which minimize water loss and maximize CO2 usage; they are known as the C4-syndrome (C4 photosynthesis), so called because CO2 is fixed to give 4-carbon compounds, and crassulacean acid metabolism (CAM).
C4 anatomy
In a typical C3 leaf, the majority of chloroplasts are distributed throughout the palisade mesophyll (below the epidermis) and the spongy mesophyll. These cells are somewhat randomly arranged between large gas-spaces that connect to the stomatal pores . In the leaf of a C4 plant, the vascular bundles are surrounded by a ring of bundle-sheath cells containing chloroplasts. These are surrounded by loosely packed mesophyll cells and air-spaces . This isknown as ‘Krantz’ anatomy (German for ‘a wreath’). The bundle-sheath and mesophyll cells are interconnected by many plasmodesmata and no bundle sheath cell is separated by more than a few cells from a mesophyll cell.
C4 biochemistry
In a C4 leaf, the first product of CO2 fixation is a 4-carbon acid. This is produced by the action of an additional cycle, the Hatch/Slack pathway, found in the mesophyll cells. The first stage of this pathway is catalyzed by the enzyme phosphoenolpyruvate (PEP) carboxylase. PEP-carboxylase uses HCO3 – (formedwhen CO2 dissolves in the cytoplasm) and PEP as substrates, yielding theC4 acid oxaloacetate which is converted to eithermalate or aspartate (both C4acids) and they are transported directly to the bundle sheath cells, where theC4 acid is decarboxylated to yield a C3-acid and CO2. At this stage, the CO2released is fixed by the Calvin cycle and the C3 acid transported back to themesophyll cell (Fig. 1). The system therefore functions as a C3 pathway to which a ‘CO2 concentrator’ has been added, transporting CO2 into the bundle sheath cells, and generating a high CO2 environment for Rubisco. The system has two major effects. First, it reduces photorespiration to undetectable levels as the Rubisco is saturated with CO2 giving conditions in which its oxygenase function is inhibited . This improves photosynthetic efficiency by up to 30%. Second, Rubisco is able to function at optimal CO2 concentrations, well above atmospheric levels. PEP-carboxylase is also saturated at the concentration of HCO3– in the cytosol which results from atmospheric CO2.
There is an energy cost to C4, however. The regeneration of the C3 acid PEP from the C3-acids transported requires the consumption of adenosine triphosphate (ATP) with the cleavage of both phosphates to yield adenosine monophosphate (AMP).
CAM anatomy
CAM plants are generally adapted to survive drought; they are succulents with fleshy leaves and few air spaces. Unlike C4 plants, they lack the physical compartmentation of photosynthesis into two cell types, but instead fix CO2 in the night as C4 acids that are stored in a vacuole which occupies much of their photosynthetic cells’ volume. The stomata of CAM plants are closed during the most desiccating periods of the day and are open at night.
CAM biochemistry
In common with C4 plants, the first stage of CO2 fixation in a CAM plant involves PEP-carboxylase, which incorporates CO2 into a C4-compound, oxaloacetate, in the dark. NADPH is used in the conversion of oxaloacetate to malate; malate is then stored in the vacuole (Fig. 2). In the light, malate is released from the vacuole and broken down to yield CO2, which is then used in the Calvin cycle. CAM photosynthesis eliminates photorespiration and increases efficiency, though losses result from the energy expended in transporting C4 acids to the vacuole and in the formation of malate. CAM plants may have a major advantage in drought conditions, with the ability to photosynthesize in daylight with closed stomata.
Distribution of CAM and C4 photosynthesis
The C4 syndrome occurs in many plant families. The syndrome is most common in arid environments. C4 plants are distributed throughout the world and C4 crops such as maize have been successfully introduced into temperate climates. C4 plants are found in a number of families (Table 1), but by no means all the members of those families are C4 species. CAM plants are similarly distributed in many families, mostly succulents and are distributed in arid zones throughout the world. Some families contain C3, CAM and C4 species. Although there appears to be an advantage to C4 and CAM, many arid zone plants are C3 and use other adaptations to survive.