Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Fishes as predators
Attack and capture - Fishes as predators
Attack and capture
An actual attack takes place as the predator launches itself at the prey and engulfs the prey in its mouth. The evolutionary advance of fishes is synonymous with the development of jaws, which function to surround, impale, or inhale prey and then pass the prey posteriorly for processing (=handling or subduing; see below). Active prey are brought into the mouth by overtaking, extending the mouth, and suction, often in combination. Fast start predators overtake or intercept their prey and impale them on sharp teeth. Overtaking may involve rapid swimming via body musculature, but many fishes also shoot the mouth out to surround the prey, as in Largemouth Bass (Nyberg 1971). The much-heralded evolution of protrusible jaws and pipette mouth, which reach their greatest development in acanthopterygian fishes, is arguably as much for overtaking prey as for suctioning.
Many benthic, lie-in-wait predators have particularly large mouths that can be opened and closed rapidly.
Frogfishes, Antennarius(Antennariidae) can expand their oral cavity up to 14-fold, engulfing prey in less than 6 ms. Stonefish, Synanceia, engulf prey in 15 ms.
Speed is not solely characteristic of large fish feeding on large prey. Zooplanktivorous damselfishes (Chromis, Pomacentridae) project their mouths to capture individual plankters in 6–10 ms; jaw protrusion followed by suction is used during capture (Coughlin & Strickler 1990). Slower movements suffi ce for nonevasive planktonic prey. Whale Sharks, Basking Sharks, manta rays, herrings, anchovies, and mackerels swim through plankton concentrations with their mouths open, passively filtering prey out of the water with their fine gill rakers. Some cyprinids and cichlids filter concentrated plankton by remaining in one place and pumping water in and out of the mouth, again using the gill rakers as a sieve (Drenner et al. 1987; Ehlinger 1989).
A few specialized predators immobilize prey before engulfing them. These specialists include torpedo rays and Electric Eels, thresher sharks, sawsharks, sawfishes, Swordfish, and billfishes. In these predators, the attack involves a high degree of energy output, followed by a relatively leisurely capture. Electricity-generating predators stun individual prey with a powerful discharge; the process
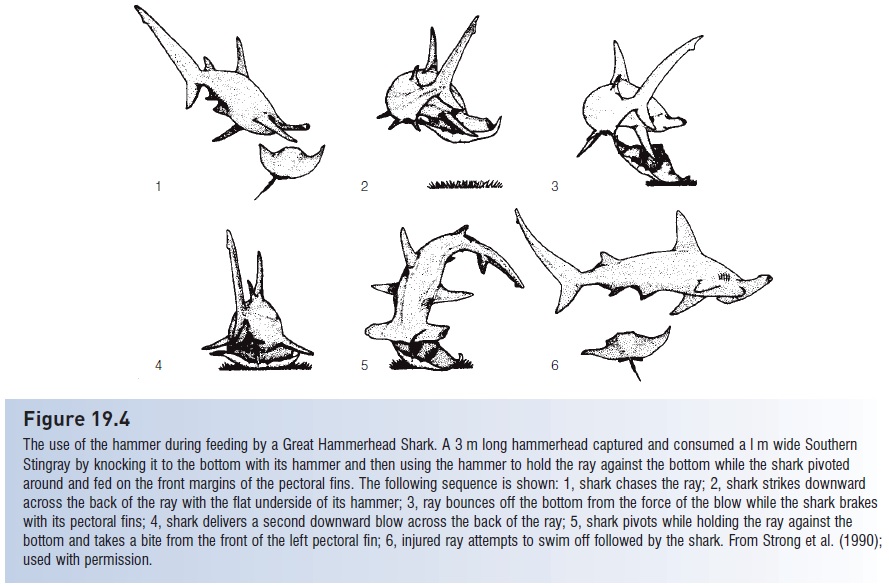
Figure 19.4
The use of the hammer during feeding by a Great Hammerhead Shark. A 3 m long hammerhead captured and consumed a l m wide Southern Stingray by knocking it to the bottom with its hammer and then using the hammer to hold the ray against the bottom while the shark pivoted around and fed on the front margins of the pectoral fins. The following sequence is shown: 1, shark chases the ray; 2, shark strikes downward across the back of the ray with the flat underside of its hammer; 3, ray bounces off the bottom from the force of the blow while the shark brakes with its pectoral fins; 4, shark delivers a second downward blow across the back of the ray; 5, shark pivots while holding the ray against the bottom and takes a bite from the front of the left pectoral fin; 6, injured ray attempts to swim off followed by the shark. From Strong et al. (1990); used with permission.
In torpedo rays, the predator encompasses the prey with its pectoral fins and discharges its electric organ. The immobilized prey is then grasped in the mouth. Thresher sharks, sawfishes, Swordfish, and billfishes are among the few predators that are minimally bothered by the confusion created by a large group of prey fishes (see Responses of aggregated prey). These predators enter a school and slash laterally with their bill (or tail in the thresher shark), and then pick up incapacitated or decapitated prey. The actual pattern is poorly understood.
Another fish that incapacitates its prey prior to capture is the archerfish (Toxotidae), which feeds on terrestrial insects. Archerfishes have a groove in the top of the mouth along which the tongue fits. The fish propels droplets of water directionally along the groove at its aerial prey. The insects fall into the water and are snapped up by the waiting archer. Laboratory trials have demonstrated that archerfishes can learn to hit targets 30–50 cm above the water, moving as fast as 20 cm/s, with up to 50% accuracy. This task requires correcting not only for refraction at the water’s surface and the arc and decelerating velocity of the fi red droplet but also the change in the three dimensional location of the target (Dill 1977a; Schuster et al. 2006). What makes this behavior even more astounding is that individuals can learn to adjust their shots by observing the efforts of shoalmates (Fig. 19.5). Osphronemid fishes in the genusColisa also spit water at insects.
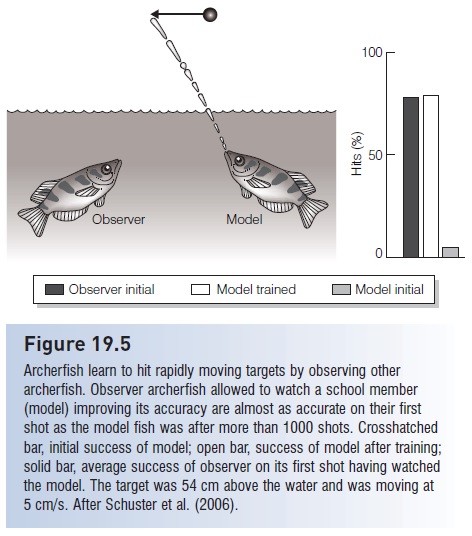
Figure 19.5
Archerfish learn to hit rapidly moving targets by observing other archerfish. Observer archerfish allowed to watch a school member (model) improving its accuracy are almost as accurate on their first shot as the model fish was after more than 1000 shots. Crosshatched bar, initial success of model; open bar, success of model after training; solid bar, average success of observer on its first shot having watched the model. The target was 54 cm above the water and was moving at 5 cm/s. After Schuster et al. (2006).
The actual strike of pikelike predators is short and fast, involving a prestrike S-shaped bending of the body and maximal forward propulsion driven by the combined surface area of the median and caudal fins; pike acceleration in this phase has been measured at more than 150 m/s (Webb 1986; Dominici & Blake 1997). Prey are impaled on the sharp jaw teeth, manipulated into a head-first position, and swallowed; in barracuda, large prey can be cut into smaller pieces for swallowing. In most predators, the attack is focused on the center of mass of the prey’s body, because the escape response involves a pivoting on the center of mass and hence the center moves least relative to other prey body parts.
When attacking prey in shoals, the major obstacle to successful prey capture is the presence of other shoal members; a direct relationship exists between likelihood of escape and numbers of individuals in the shoal. Hence many predators engage in tactics that separate prey individuals from the group. For example, when Pike (Esox lucius, Esocidae) attack minnow (Phoxinus phoxinus) schools, the predator’s strike often leaves individual prey separated from the school. The predator then preferentially chases these individuals, which may account for 89% of the predator’s success (Magurran & Pitcher 1987). Similar group-separating tactics are employed by other predators, such as Blue Jack, Caranx melampygus (Carangidae), when attacking mixed species schools of snappers, and piranha, Serrasalmus spilopleura (Characidae), attacking cichlid shoals (Potts 1980; Sazima & Machado 1990). Stragglers are up to 50 times more likely to be attacked than fish within a group, and success rates for attacks on stragglers can be four times higher than for attacks on the main shoal.
Aggressive mimicry also occurs in a group context, the victims either being fish and invertebrates exterior to the shoal or even unsuspecting members of the shoal itself. In both cases, shoal membership by the mimic allows the predator to get close enough to attack. Juveniles of the Indo-Pacific grouper, Anyperodon leucogrammicus (Serranidae), swim with and resemble adult females of four similar species of wrasses (Labridae). Small fishes have no reason to fear the wrasses, which feed on a variety of small benthic invertebrates. However, the grouper is a piscivore and has been observed snaring small damselfishes while swimming among the shoaling wrasses. Similarly, hamlets of the West Indian genus Hypoplectrus (Serranidae) resemble damselfishes and angelfishes, the models here being zooplanktivores or herbivores. The hamlets are carnivores and their presumed mimicry could allow them to sneak up on invertebrate prey that do not distinguish them from the otherwise harmless, and usually more numerous, model species (Moland et al. 2005; Randall 2005). Some scaleeating fishes attack from within shoals, their primary prey being shoalmates that they resemble. These predators include a characid, Probolodus heterostomus, which feeds on schooling characids in the genus Astyanax, and a cichlid, Corematodus shiranus, which resembles and schools with
Prey choice is also affected by relative numbers of different kinds of individuals in the prey shoal. Oddity in appearance or behavior often stimulates attack. In mixed schools, the minority species is often attacked disproportionately, as happens when Gafftopsail Pompano (Carangidae) feed on anchovetas (Engraulidae) that are schooling with more numerous Flatiron Herring (Clupeidae) (Hobson 1968). In experimental trials, Largemouth Bass are much more successful predators when one or two blue-dyed minnows are added to small minnow shoals; the odd individuals are taken preferentially (Landeau & Terborgh 1986). Predation on odd individuals within a school may result from them standing out against the background of the more common type or, as suggested by Landeau and Terborgh (1986), because oddity alleviates the confusion effect created by a mass of similar-appearing prey animals. Whether predatory fishes are more likely to attack injured individuals within a group remains unclear. Certainly sharks and barracudas are attracted to injured or erratically swimming prey, and the literature on mammalian predators (e.g., lions, wolves, hyenas) suggests that fishes would also feed preferentially on injured individuals. However, the only quantitative study of the subject indicates, if anything, an avoidance of injured prey (Major 1979). Regardless, predation on odd individuals indicates the strong selection that occurs for uniformity of appearance and behavior within a shoaling species.
Related Topics