Chapter: Essentials of Anatomy and Physiology: Lymphatic System and Immunity
Activation and Multiplication of Lymphocytes
Activation and Multiplication of Lymphocytes
The specialized B-cell or T-cell clones can respond to antigens and produce an adaptive immune response. For the adaptive immune response to be effective, two events must occur: (1) antigen rec-ognition by lymphocytes and (2) proliferation of the lymphocytes recognizing the antigen.
Antigen Recognition
Lymphocytes have proteins, called antigen receptors, on their surfaces. The antigen receptors on B cells are called B-cell receptors, and those on T cells are called T-cell receptors. Each receptor binds with only a specific antigen. Each clone consists of lympho-cytes that have identical antigen receptors on their surfaces.
When antigens combine with the antigen receptors of a clone, the lym-phocytes in that clone can be activated, and the adaptive immune response begins.
B cells and T cells typically recognize antigens after large mol- ecules have been processed or broken down into smaller fragments Antigen-presenting cells, such as macrophages, present antigens to B and T cells. The antigens are taken into macrophages by phago cytosis and are broken down into smaller antigen fragments. The processed antigen fragments are bound to major histocompatibility complex molecules, transported to the surface of the macrophages, and presented to B cells and T cells (figure 14.10, step 1).
Major histocompatibility complex (MHC) molecules are glycoproteins that have binding sites for antigens. Different MHC molecules have different binding sites—that is, they are specific for certain antigens. It is important not to confuse the MHC molecules with the antigen receptors described above. Though both types of receptors interact with antigens, MHC molecules are a different group of receptors found on the membrane of many types of cells. There are two classes of MHC molecules. MHC class I molecules are found on the membranes of most nucleated cells and MHC class II molecules are found on the membranes of antigen-presenting cells, B lymphocytes, and other defense cells. The MHC molecules function as “serving trays” that hold and present a processed antigen on the outer surface of the cell membrane. The combined MHC molecule and processed antigen can then bind to the antigen recep- tor on a B cell or T cell and stimulate it. For example, figure 14.10, step 2, illustrates how helper T cells are stimulated when combined with MHC class II molecules.
The MHC molecule/antigen combination is usually only the first signal necessary to produce a response from a B cell or T cell.
In many cases, costimulation by a second signal is also required. Costimulation can be achieved by cytokines (sı̄′ tō-kı̄nz), which are proteins or peptides secreted by one cell as a regulator of neighboring cells. For example, interleukin-1 (in-ter-loo′ kin) is a cytokine released by macrophages that can stimulate helper T cells (figure 14.10, step 3).
Lymphocytes have other surface molecules besides MHC molecules that help bind cells together and stimulate a response. For example, helper T cells have a glycoprotein called CD4, which helps connect helper T cells to the macrophage by binding to MHC class II molecules. The CD4 protein is also bound by the virus that causes AIDS (see the Diseases and Disorders table). As a result, the virus preferentially infects helper T cells. Cytotoxic T cells have a glycoprotein called CD8, which helps con-nect cytotoxic T cells to cells displaying MHC class I molecules.
Lymphocyte Proliferation
Before exposure to a particular antigen, the number of helper T cells that can respond to that antigen is too small to produce an effective response against it. After the antigen is processed and presented to a helper T cell by a macrophage, the helper T cell responds by producing interleukin-2 and interleukin-2 receptors(figure 14.10, step 4). Interleukin-2 binds to the receptors and stimulates the helper T cell to divide (figure 14.10, step 5). The “daughter” helper T cells produced by this division can again be presented with the antigen by macrophages and again be stimulated to divide. Thus, the number of helper T cells is greatly increased (figure 14.10, step 6).
It is important for the number of helper T cells to increase because helper T cells are necessary for the activation of most B cells or T cells (figure 14.10, step 7). For example, B cells have receptors that can recognize antigens. Most B cells, however, do not respond to antigens without stimulation from helper T cells. Without functional helper T cells, the immune response of B cells would not be effective to prevent disease.
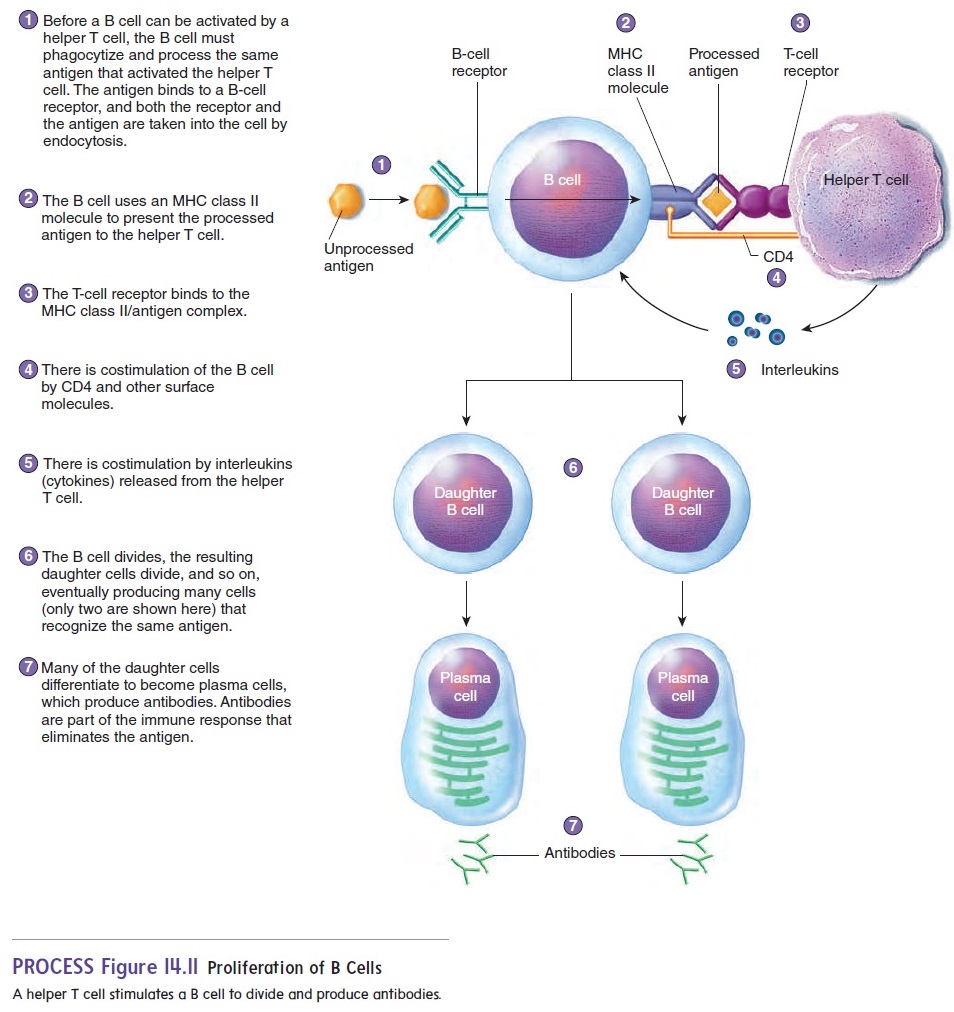
B-cell proliferation begins when a B cell takes in the same kind of antigen that stimulated the helper T cell (figure 14.11, step 1). The antigen is processed by the B cell and presented on the B-cell surface by an MHC class II molecule (figure 14.11, step 2). A helper T cell is stimulated when it binds to the MHC class II/antigen complex (figure 14.11, step 3). There is also costimulation involving CD4 and interleukins (figure 14.11, steps 4 and 5). As a result, the B cell divides into two “daughter” cells (figure 14.11, step 6). These daughter cells may differentiate into plasma cells, which produce antibodies (figure 14.11, step 7).The division process continues, eventually producing many cells capable of producing antibodies and resulting in sufficient anti-bodies to destroy all the antigen.
Related Topics