Chapter: Plant Biology : Vegetative anatomy
Vegetative anatomy of Herbaceous stems and primary growth
HERBACEOUS STEMS AND PRIMARY GROWTH
Key Notes
Shoot structure
Herbaceous stems are green and non-woody. They have a range of growth forms generated by an apical meristem that produces leaf and bud primordia. The shoot is surrounded by an epidermis, within which lies the cortex. Vascular tissues occur as a ring of separate bundles towards the outside of the stem in dicotyledons and in a scattered pattern
in monocotyledons. The center of the stem is the pith parenchyma.
Meristems
The shoot meristem has a tunica producing the epidermis and the layers of cells beneath it while the corpus produces the cortex, pith and vascular tissues. The meristem can also be divided into the central zone where cell division occurs, the peripheral zone where leaves, shoots and new meristems form, and the rib zone where stem growth occurs.
Vascular tissue
Vascular bundles containing phloem and xylem form a ring or a complex array throughout the inner part of the stem. In some dicotyledons the ring is almost continuous.
Architecture
The shape of the stems is governed by the controlled positioning of leaves and buds by the meristem. Their positioning may be spiral, distichous, opposite, decussate or whorled.
Shoot structure
Herbaceous stems are green throughout without deposition of lignin that would make them woody. Shoots are more complex than roots and take on a wide range of growth forms reflecting function. Like the root, the shoot has an apical meristem. Unlike the root (where lateral branching only occurs in mature tissue), the shoot meristem produces lateral shoots and leaves (Fig. 1).
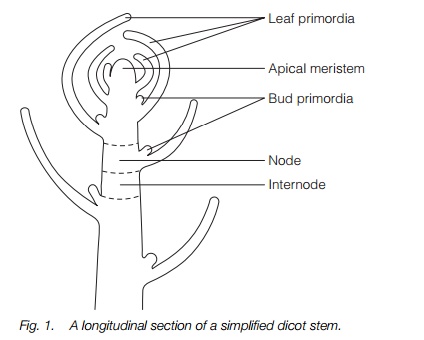
The apical meristem produces leaf primordia (which will form leaves) and bud primordia (which will form shoots). These are produced in position and order which gives rise to the characteristic form of the shoot which is recognizable for each species (see ‘architecture’ below).
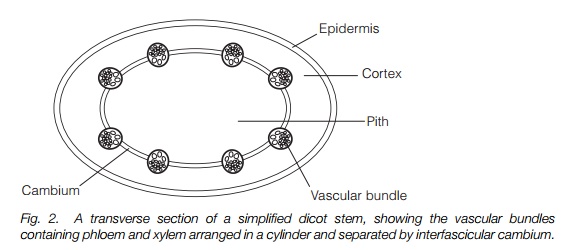
The shoot is surrounded by an epidermis. This outer layer provides the protective barrier between the stem and its environment and is covered in a lipid-based protective substance, cutin (Topics I1 and M4). Within the epidermis, cells of the ground tissues, the cortex, may be photosynthetic and occupy the space surrounding the vascular bundles. In some species, the cortex is only a few cell layers, in others many more. Within the cortex, bounded by the vascular bundles, the center of the stem is occupied by pith (Fig. 2).
Meristems
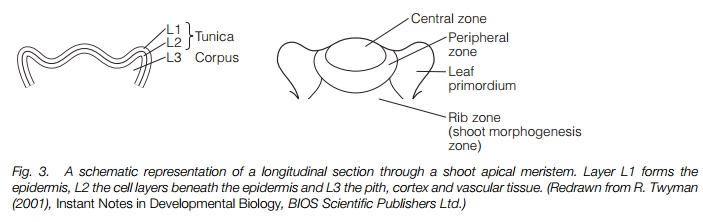
The shoot meristem produces the cells that will form the major tissues of new stems and leaves. It is organized in layers of cells each producing a different tissue and in zones with different functions (Fig. 3). Layer 1 produces the epidermis that surrounds the shoot. Here cells divide to generate only a single layer. Beneath it, layer 2 divides to generate the layers of cells beneath the epidermis. Layer 1 and layer 2 together are known as thetunica. Layer 3 produces all the other tissues of the shoot: the cortex, pith and vascular tissues, and is known as the corpus. The central zonecontains actively dividing cells. Leaves and shoots (including new meristems) form in the peripheral zone. Stemgrowth occurs in the rib zone and cells mature to form the tissues of the shoot.
Vascular tissue
The vascular tissue of the shoot commonly occurs as a ring of separate vascular bundles, each containing phloem and xylem. They are located between the cortex and pith (Fig. 2) in dicotyledons, while in monocotyledons they are found in a scattered array throughout the inner part of the stem. All vascular bundles contain phloem and xylem strands in parallel to one another. Associated with the xylem strands are xylem parenchyma cells and fibrous sclerenchyma tissue may be associated with the phloem. In some dicotyledons, the vascular bundles form an almost continuous ring; however, individual vascular bundles remain, separated by narrow bands of interfascicular parenchyma (the site of origin of the cambium for secondary growth) which separate the cortex from the pith (Fig. 2).
Architecture
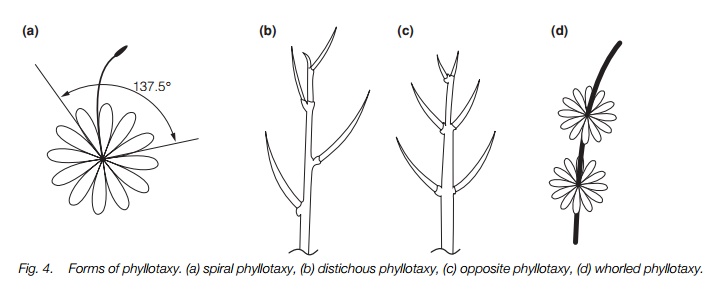
The shape of the stems of different species are clearly recognizable from one another (Fig. 4). For instance, before flowering, Arabidopsis thaliana(Topic E1) forms a rosette of leaves, each initiated 137.5° from the preceding one formed. This process, termed spiral (or helical) phyllotaxy, is common in many species and results in the formation of a spiral pattern of growth. In other species, new leaves may be formed singly at each node in two opposite rows (distichousphyllotaxy), in pairs opposite one another (opposite phyllotaxy) or in whorls of five or more at one node (whorled phyllotaxy). Where pairs of leaves form at right angles to a previous pair of leaves, the plant has decussate phyllotaxy.
Related Topics