Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Conservation and the future of fishes
Species introductions - General causes of fish biodiversity decline
Species introductions
Movement of species into new areas is a natural zoogeographic phenomenon. When such range extension occurs as a result of human actions, it is considered anintroduction. Natural dispersal is limited by a species’ mobility and by physical barriers. Under natural conditions, species are constrained by co-evolutionary processes; species have natural parasites, predators, and competitors that control population growth, and organisms typically exploit prey taxa that have evolved defense mechanisms against the predator’s foraging tactics. When individuals of a species are introduced suddenly into an alien environment, they may find the new physical and biological factors inhospitable or even lethal. Alternatively, freedom from natural biotic control may remove all checks on population growth. It is these liberated aliens that cause the greatest problems. Many such introductions have become well-known pests: rabbits, cane toads, and prickly pear cactus in Australia;
starlings, English sparrows, gypsy moths, and zebra mussels in North America; mongoose and mynah birds in Hawaii; feral goats in the Galápagos and on many other islands, to name a few. These catastrophic introductions have their counterparts in fish assemblages as well.
Introductions go by a great variety of names, including alien, allochthonous, exotic, feral, introduced, invasive, naturalized, nonindigenous, non-native, transplanted, and translocated. Offi cial US terminology under the Nonindigenous Aquatic Nuisance Prevention and Control Act of 1990 refers to nonindigenous organisms, but increasingly acceptance is leaning to simply alien (see Lever 1996, and especially Fuller et al. 1999). Sometimes a distinction is made between transplants that are moved within their country of origin but outside their native range, versus exotic species that are introduced into a new country. In terms of ecological impact, such geopolitical distinctions are meaningless (Fuller et al. 1999). Introductions may occur through deliberate actions (gamefish stocking, vegetation control, aquaculture, aquarium releases) or inadvertent mishaps (ballast water introductions, aquaculture escapement, bait fish release).
Untold hundreds of species of fishes have been deliberately transported among different countries. In the USA alone, approximately 536 alien fish taxa (species, hybrids, and unidentified forms) have been introduced, 35% imported from foreign countries and 61% translocated within the nation (Fuller et al. 1999; Nico & Fuller 1999; http://nas.er.usgs.gov). Half of the foreign exotics have established breeding populations. Most of these fishes represent deliberate introductions by government agencies and individuals (e.g., Grass Carp, Ctenopharyngodon idella, for vegetation control; Peacock Cichlid, Cichla ocellaris, as a
Table 26.2
Ten commonly introduced but controversial fish species. Species listed have often been effective in terms of the original purpose for which they were introduced, but have subsequently posed serious ecological problems. From Helfman (2007), based on Courtenay et al. (1984), Welcomme (1984, 1988), Lever (1996), and Fuller et al. (1999). The number of countries and island groups where established is from Lever (1996), the number of US states where introduced is from Fuller et al. (1999). Adapted from Helfman et al. (1997), and presented in roughly phylogenetic order.
Depending on one’s perspective, the same introduction can be claimed as a success story or reviled as a disaster (Fuller et al. 1999) (Table 26.2). Eight fish species are included among “100 of the world’s worst invasive species” as recognized by the Global Invasive Species Database of the IUCN’s Invasive Species Specialist Group. These eight are the Walking Catfish, Clarias batrachus; Common Carp; Rainbow and Brown trout; Mosquitofish; Largemouth Bass; Nile Perch; and Mozambique Tilapia (see Table 26.2 for details on most of these). Many of these species have been spread around the world and their ecological impacts vary from minor to catastrophic (Tilapia supply significant protein to humans in need, although alternative, edible native species exist, or existed, in many locales). These fishes are listed by IUCN alongside such known bad actors as chestnut blight, water hyacinth, prickly pear cactus, kudzu, fi re ants, malaria mosquitoes, gypsy moths, zebra mussels, cane toads, brown tree snakes, starlings, rats, rabbits, pigs, and goats.
One outcome of this enthusiasm for moving fishes around the globe is homogenization of previously unique assemblages that evolved at particular locales. As can be seen in Table 26.2, the same fishes have been introduced repeatedly in different places: six of the 10 listed species have been introduced into 40 or more different countries. Not coincidentally, considerable overlap exists among the receiving nations, hence most countries contain established populations of several of the species listed. Introductions are often most successful in degraded habitats that are no longer suitable for local endemics, which means that alien species are replacing and displacing native species. When we total up the introductions and losses, we find that widely separated locales have similar faunal lists that are dominated by alien species. This repeated observation, which includes not just fishes but birds, mammals, insects, and plants, amounts to homogenization of the earth’s
fauna and flora: everywhere looks the same (Lockwood & McKinney 2001). The process is so widespread, its outcome so ubiquitous, and the disruption so thorough that Williams and Meffe (1998, p. 118) declared that, “The continued homogenization of the world’s flora and fauna . . . is an ecological holocaust of major proportions”. Gordon Orians of the University of Washington refers to the era in which we live as the Homogocene.
Homogenization of fish faunas is especially prevalent in temperate, developed nations where introduction “efforts” have been ongoing. In Europe, Cowx (1997) listed 166 introduced or translocated fishes, which amounts to about 46% of Europe’s 358 native species. Fifteen species have been introduced into 10 or more countries, including Carp (24 countries), Brook Trout (23), Grass Carp (20), Pumpkinseed Sunfish (19), and Rainbow Trout (nine). Other countries with low native diversity or high numbers of introduced species – where extensive homogenization would be expected and where its impacts should be monitored – are New Zealand, Australia, and South Africa.
Homogenization of US fish faunas has happened on a grand scale (Fuller et al. 1999; Rahel 2000, 2002). Of the 76 species introduced into 10 or more states, 32 have been placed in 25 or more states and 13 have gone into more than 35 states. Eight of the latter 13 are relatively large piscivores that continue to be stocked as game species in many places. Three (Common Carp, Goldfish, and Tench) stocked extensively during the late 19th century are now generally regarded as nuisance species.
How have stocking efforts affected fish diversity? Rahel (2000) compared historical lists with current species lists and found that state faunas have grown significantly more similar over time. In the past two centuries, similarity among fish faunas of states has increased by an average of 15 species, with almost 20% of states sharing 25 or more additional species. Over half of the fish faunas of Nevada, Utah, and Arizona are non-native, containing species brought in primarily from the eastern part of the country to improve angling. At the same time, and in combination with the habitat degradation that makes regions hospitable to aliens and inhospitable to local endemics, the list of imperiled US fishes has grown.
Homogenization is generally discussed with regard to exotics displacing natives, but the process also involves movement of widespread, generalist native species into areas or habitats previously occupied by local endemics. As is occurring in the southern Appalachian mountains of the USA, localized endemics adapted to clear, cool, low-productivity streams in upland regions are progressively replaced by widespread, generalist species more common in lower, more productive portions of river networks (Scott & Helfman 2001) (Fig. 26.6). The factors responsible for this replacement scenario include upland and riparian deforestation. Subsequent erosion of the uplands causes infilling of rapid and riffl e habitats due to increased sediment loads; streams also become broader, deeper, and warmer. Cool water, endemic specialists in shallow, fastfl owing habitats (darters, sculpins, benthic minnows) arereplaced by warm water generalists that can live in a variety of habitats but especially in slower flowing habitats (sunfishes, pool-dwelling minnows, suckers) (Jones et al. 1999a, Walters et al. 2003). Because the endemics were localized and the generalists were widespread, faunas of different drainage basins increase in similarity. Habitat homogenization promotes biotic homogenization (e.g., Boet et al. 1999; Marchetti et al. 2001), and homogenization occurs even though almost all species involved are technically native to the area.
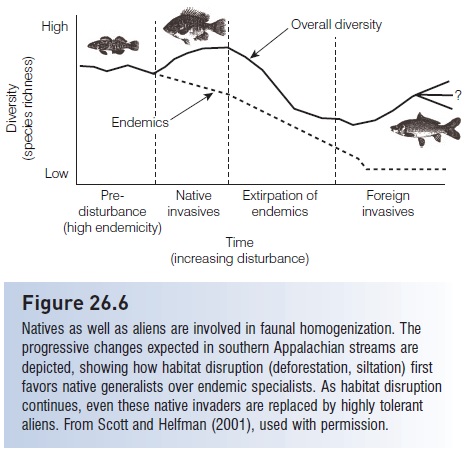
Figure 26.6
Natives as well as aliens are involved in faunal homogenization. The progressive changes expected in southern Appalachian streams are depicted, showing how habitat disruption (deforestation, siltation) first favors native generalists over endemic specialists. As habitat disruption continues, even these native invaders are replaced by highly tolerant aliens. From Scott and Helfman (2001), used with permission.
Introduced predators
What havoc the introduction of any new beast of prey must cause in a country, before the instincts of the indigenous inhabitants have become adapted to the stranger’s craft or power. Charles R. Darwin (1871) Introductions can lead to population reduction or extermination of native fishes, either directly through predation on adults, eggs, and young, or indirectly through superior competition, hybridization, or transmission of pathogens (Balon & Bruton 1986; Fausch 1988; Ross 1991). Some catastrophic introductions are inadvertent, as with the spread of the Marine Lamprey, Petromyzon marinus, into the North American Great Lakes probably via manmade canals; Lake Trout, whitefishes, pike-perch, and other species declined precipitously in the wake of the lamprey (Daniels 2001; Petromyzontiforms). Predatory species that have been widely introduced to provide sportfishing are the Peacock Cichlid (Cichla ocellaris), Largemouth Bass (Micropterus salmoides), and Rainbow and Brown trout (Oncorhynchus mykiss and Salmo trutta).
Such introductions often decimate native fish faunas, including reduction of important food fishes. Peacock Cichlids escaped from an impoundment and into the Chagres River, Panama. The cichlid invaded Gatun Lake and progressively eliminated seven local fish species, including an atherinid, four characins, and two poeciliids; vegetation increased and fish-eating birds were displaced (Zaret & Paine 1973; Swartzmann & Zaret 1983).
Largemouth Bass have been responsible for similar community disruptions in Lago de Patzcuaro, Mexico; in Lake Naivasha in Kenya; in northern Italy; in Zimbabwe and South Africa; and in Lake Lanao, Philippines. Rainbow and Brown trout have led to the decline of endemic fishes in Yugoslavia, Lesotho, Colombia, Australia, New Zealand, South Africa, and in Lake Titicaca in Bolivia and Peru (McDowall 2006). In Lake Titicaca, the world’s highest lake, a species flock of numerous cyprinodontids (Orestias) has been decimated, first through direct predation and later via competition for invertebrate prey. Brown Trout in particular have been identified as an effective predator on native fishes, including other salmonids. Brown Trout have contributed to the decline of several threatened salmonids, including Gila Trout (Oncorhynchus gilae), McCloud River Dolly Varden (Salvelinus malma), and Golden Trout (Oncorhynchus mykiss aguabonita), the latter being the offi cial state fish of California. Introduced salmonids have been particularly destructive to galaxioid fishes through much of the geographic range of this group of southern hemisphere fishes (McDowall 2006).
One of the most dramatic examples of the effects of an introduced predator involves the stocking of the latid Nile Perch, Lates cf. niloticus, in Lakes Victoria and Kyoga, east Africa (Ogutu-Ohwayo 1990; Kaufman 1992; Witte et al. 1992; Lowe-McConnell 1997; Ogutu-Ohwayo et al. 1997); debate rages over exactly which Lates species was introduced and how often (Pringle 2005). Lake Victoria is, or was, a showcase of evolution and explosive speciation among fishes, having given rise to a species flockof perhaps 300 haplochromine cichlids, as well as three dozen other fishes. The lake is thought by many to have contained the richest lacustrine fish fauna in the world. Against the advice of ecological experts, Nile Perch were stocked in the lakes in the early 1960s, “to feed on ‘trash’ haplochromines . . . [and convert them] into more desirable table fish” (Ribbink 1987, p. 9).
This predator, which can attain a length of 2 m and a weight of 200 kg, spread slowly through both lakes, effectively wiping out native fishes by feeding preferentially on abundant species, then shifting to other species as the density of the initial prey declined, and finally turning to cannibalism. Commercial landings of cichlids went from 27 to 0 kg/h and fell from 32% to 1% of the catch between 1977 and the early 1980s. Nile Perch landings increased to 169 kg/h (Seehausen et al. 1997b). As species were eliminated, food webs in the lakes were substantially disrupted and simplified (Fig. 26.7); elimination of herbivorous cichlids led to algal blooms and attendant oxygen depletion in deep water, which caused periodic fish kills. In Lake Kyoga, the catch changed from a multispecies fishery dominated by several haplochromines to one dominated by two introduced species (Nile Perch and a tilapia) and a native cyprinid (Rastrineobola argentea).
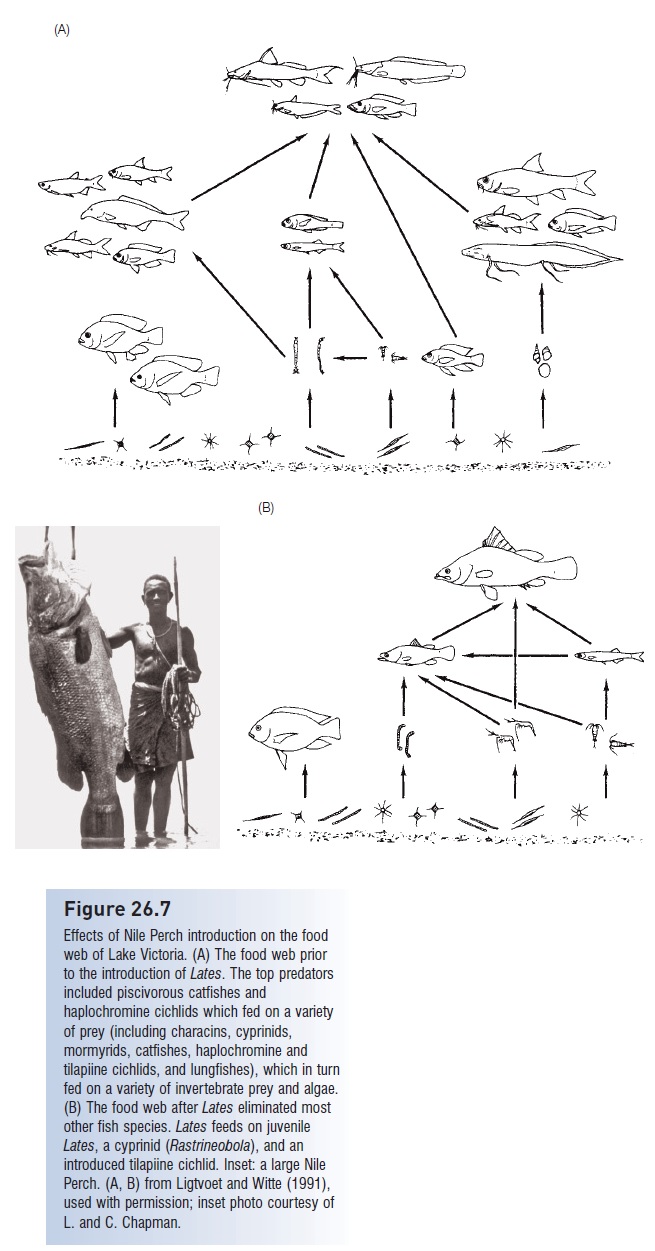
Figure 26.7
Effects of Nile Perch introduction on the food web of Lake Victoria. (A) The food web prior to the introduction of Lates. The top predators included piscivorous catfishes and haplochromine cichlids which fed on a variety of prey (including characins, cyprinids, mormyrids, catfishes, haplochromine and tilapiine cichlids, and lungfishes), which in turn fed on a variety of invertebrate prey and algae. (B) The food web after Lates eliminated most other fish species. Lates feeds on juvenile Lates, a cyprinid (Rastrineobola), and an introduced tilapiine cichlid. Inset: a large Nile Perch. (A, B) from Ligtvoet and Witte (1991), used with permission; inset photo courtesy of L. and C. Chapman
Just how many of the endemic cichlids have actually been exterminated is difficult to say: perhaps only 50% of the species have been described and rare fishes are difficult to sample (Goldschmidt 1996). However, decreasing catches indicate that populations are shrinking and the continued threat of predation by Nile Perch and commercial fishing will only exacerbate the situation. Based on comparative samples taken in 1978 and 1990, approximately 70%, or 200 species, of haplochromines are extinct or threatened with extinction (Witte et al. 1992). Given present trends, “probably more vertebrate species are at imminent risk of extinction in the African lakes than anywhere else in the world” . Events in Lake Victoria call into question recent proposals for introduction of Nile Perch into Lakes Malawi and Tanganyika and point out the ecological consequences of introducing predators into any aquatic system (Witte et al. 1992).
The scenario played out in Lakes Victoria and Kyoga is one of reduced biodiversity and simplified community interactions as a cost of production of animal protein for human consumption. Successful fisheries for introduced Nile Perch and Tilapia have been established in those lakes, replacing the previous fisheries for smaller, native haplochromines. The impacts of these introductions are not, however, limited to the aquatic ecosystems. Nile Perch have a relatively high oil content. Traditional preparation methods, such as air drying, are less effective for processing Nile Perch. Instead, the flesh is often smoked over wood fi res, which leads to deforestation of hillsides in the Lake Victoria basin, runoff of sediment and nutrients into the lake, reduced light transmission affecting reproduction of cichlids, and eutrophication leading to deoxygenation of deeper waters, to which Nile Perch are intolerant (Seehausen et al. 1997a, 1997b; Kitchell et al. 1997); low oxygen areas may serve as refuges for native cichlids (Schofield & Chapman 2000). An unexpected result of combined ecological and socioeconomic influences is that condition factors of Nile Perch in Lake Victoria have declined to the lowest values known for the species anywhere (Ogutu- Ohwayo 1999). Similar introductions, for similar purposes and with similar results, could be cited. For example, Contreras and Escalante (1984) identified nine instances in Mexico where, after the introduction of potential food fishes, the number of native, often endemic species declined by an average of 80%.
Competition
Predation of new fishes on old is the most obvious effect of introduced species. Less well documented, but of potentially serious consequence, is the threat of competition, disease, and hybridization that can occur from introducing foreign species (Taylor et al. 1984). Competition is difficult to prove even under the best controlled, experimental conditions. Evidence of competitive depression of native fishes usually takes an inferential form, in terms of overlap in use of potentially limiting resources, or decline in native fishes correlated with the introduction of a nonpredator. Diet overlap with native fishes in North America has been documented for such introduced species as Brown Trout, Common Carp, Pike Killifish (Belonesox belizanus), numerous cichlids, and two Asiatic gobies (Acanthogobius flavimanus and Tridentiger trigonocephalus). Blue Tilapia (Oreochromis aureus) overlap extensively in diet with Gizzard Shad and Threadfin Shad (Dorosoma cepedianum and D. petenense, Clupeidae). Blue Tilapia reproduce rapidly, forming dense populations (>2000 kg/ha) of stunted individuals. Introductions of Blue Tilapia in Texas and Florida have resulted in concomitant population declines of shad, particularly of the benthicfeeding Gizzard Shad. Overcrowding by tilapia also inhibits Largemouth Bass spawning behavior, although the actual mechanisms involved (competition, chemical suppression, behavioral interference) are poorly understood (Taylor et al. 1984). Competition for food probably explains the negative impact of introduced Guppies (Poecilia reticulata)
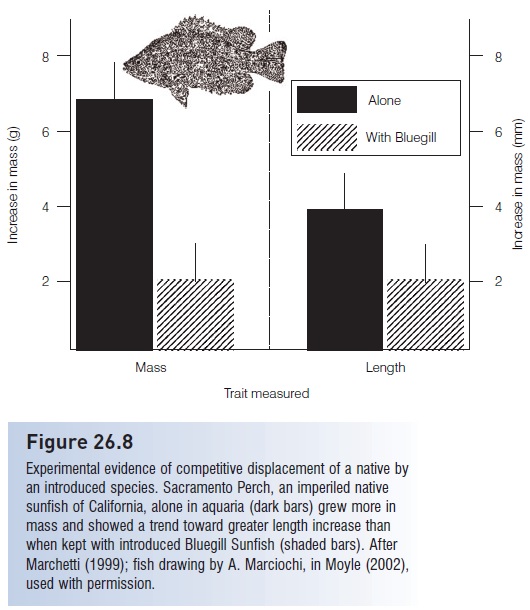
Figure 26.8
Experimental evidence of competitive displacement of a native by an introduced species. Sacramento Perch, an imperiled native sunfish of California, alone in aquaria (dark bars) grew more in mass and showed a trend toward greater length increase than when kept with introduced Bluegill Sunfish (shaded bars). After Marchetti (1999); fish drawing by A. Marciochi, in Moyle (2002), used with permission.
Competition for nursery grounds led to a decline in catches of native Tilapia variabilis after transplantation of Redbelly Tilapia to Lake Victoria. Competitive impacts on rare, native Atlantic Salmon, Salmo salar, from abundant, escaped and released, cultured fish are a genuine concern (Jonsson & Jonsson 2006).
Our understanding of how aliens displace natives can be improved via laboratory manipulations of species and resources (e.g., Fausch 1988; Ross 1991). Marchetti (1999) looked for competitive interactions as a cause of population declines and extirpations of Sacramento Perch, Archoplites interruptus, a native California centrarchid. Sacramento Perch are least abundant where introduced sunfishes are most numerous. In lab aquaria, Marchetti found that Sacramento Perch placed with Bluegill Sunfish,Lepomis macrochirus, grew less and shifted habitat use to less natural habitats. Bluegill fed more actively and harassed the perch (Fig. 26.8).
Hybridization
Hybridization and introgression (crossing of hybrid offspring with parental genotypes) has caused rapid losses of native fishes over extensive geographic areas (Echelle 1991). Hybridization can result from habitat alterations that reduce physical and behavioral barriers between populations (e.g., Lake Victoria cichlids; Seehausen et al. 1997a). Hybridization also occurs when numbers of one species fall to the point where conspecifics are rare during mating periods, leading to interspecific matings (Hubbs 1955). Rare species have fallen victim to hybridization in the US southwest, including hybridization between the threatened Clear Creek Gambusia (Gambusia heterochir) and introduced Mosquitofish (G. affinis), between the endangered Humpback Chub (Gila cypha) and the more common Roundtail Chub (G. robusta), and between June Suckers (Chasmistes l. liorus) and Utah Suckers (Catostomus ardens). June Suckers were at one time exceedingly abundant. As their numbers decreased, in part because of water drawdowns for irrigation, June Suckers became increasingly vulnerable to genetic disruption via hybridization with abundant Utah Suckers (Echelle 1991).
Human-caused hybridization is particularly threatening where stocking programs bring hatchery or other strains of fishes into contact with native conspecifics (Utter & Epifanio 2002). Native strains disappear as they interbreed with introduced fishes, as has happened when Rainbow Trout were stocked with threatened Cutthroat, Gila, and Apache trouts (Oncorhynchus clarki ssp., O. gilae, and O. apache) in western North America (Echelle 1991). Hatchery fishes often originate from a limited gene pool or from inbred lines and have reduced genetic variability compared with wild populations. Low genetic variability correlates with lower fecundity, poorer survivorship, and slower growth, as found in different populations of endangered Sonoran topminnows (Poeciliopsis occidentalis ssp.; Quattro & Vrijenhoek 1989).
When hatchery transplants breed with wild fish, resulting offspring will often be less diverse genetically than the wild strains. Hybrid offspring may continue to breed with and eventually eliminate native stocks, as has occurred with Rainbow Trout stocked widely throughout North America. Threats from transplanted and cultured fish have caused considerable concern over the genetic integrity of wild Atlantic Salmon (Salmo salar) stocks, prompting programs to minimize the effects of sea ranching and stock enhancement programs (NASCO 1991; NRC 2004a). Even the genetic integrity of Common Carp, Cyprinus carpio, is threatened by introgressive hybridization with introduced, cultured strains. Very few wild-type carp remain in the native habitat of this widely introduced species, and the wild genetic strain has endangered status in such large river systems as the Danube (Balon 1995).
The creation of intergeneric hybrids, initially considered unlikely, has also proven troublesome. European Brown Trout (Salmo trutta) hybridize with North American Brook Trout (Salvelinus fontinalis), producing a cross known as the tiger trout (e.g., Brown 1966). The widely introduced European Rudd, Scardinius erythrophthalmus(Cyprinidae), is a hardy, colorful baitfish cultured in the southern United States. Rudd hybridize with native Golden Shiner, Notemigonus chrysoleucas. Rudd are known to be established in eight states and could potentially hybridize with Golden Shiners in 26 states in the Mississippi River basin, with unknown consequences for the fish assemblages or ecosystems of those areas. Such risks are unnecessary given that several acceptable native bait species, including the Golden Shiner, already exist throughout the region (Burkhead & Williams 1991).
Parasites and diseases
A major threat from introductions, whether exotic or transplanted, is transmission of bacterial and viral diseases and parasites to which native fishes were previously unexposed (Hedrick 1996). Furunculosis, a fatal bacterial disease caused by Aeromonas salmonicida, was originally endemic to western North American strains of Rainbow Trout. When the trout was introduced into Europe, the disease became widespread among Brown Trout populations and now occurs wherever salmonids are cultured (Bernoth et al. 1997). Whirling disease, caused by the protozoan Myxosoma cerebralis, is native and originally nonpathogenic to European salmonids (Hedrick et al. 1999). It causes swimming in tight circles, followed by postural collapse and immobility. The parasite infl ames cerebrospinal fluid, deforms the brainstem, and causes degeneration of nerves connecting the medulla and spinal cord (Rose et al. 2000). Whirling disease was transmitted from Europe to North America in the late 1950s, has proved extremely pathogenic to Rainbow and Brook trout, and is considered the single greatest threat to many US wild trout populations (MWDTF 1996). It has subsequently spread with exportation of North American salmonids, including back to Europe, where it has increased in pathogenicity. “Ich”, a debilitating gill and skin infestation caused by the ciliated protozoan Ichthyopthirius multifiliis, originated in Asia and has spread throughout temperate regions via introductions (Hoffman & Schubert 1984; Welcomme 1984; Dickerson & Clark 1998).
An interaction between genetic disease resistance and the dangers of transplantations is exemplified by fall Chinook Salmon, Oncorhynchus tshawytscha. Salmon raised from eggs taken from streams where the protozoan Ceratomyxa shasta is endemic show mortality rates of less than 14% when exposed to the pathogen. Salmon taken from streams where the pathogen is not native exhibit mortality rates of 88–100% upon exposure (Winton et al. 1983). The introduction of infected fishes into areas where specific diseases do not occur naturally, such as might occur during pen-rearing operations or a “supplementation program”, could have catastrophic consequences for endemic stocks of fishes.
Infestation problems involving native parasites can also be aggravated. Juvenile Pacific salmon (Pink and Chum, O. gorbuscha and O. keta) migrating past net pens holding Atlantic Salmon suffered mortality rates of 9–97% as a result of infestations of external copepod parasites. Infestation rates were significantly lower on juveniles that did not swim past net pen operations (Krkosek et al. 2006, 2007). Copepods (sea lice) on adult salmon are seldom fatal (see Mutualism and commensalism), but a single copepod can kill a juvenile salmon.
Ballast water introductions
A significant source of introductions receiving increasing attention is the ballast water of large ships (NRC 1996a). Water is pumped into special ballast tanks or empty holds of ships to stabilize them; this water is then pumped out when cargo is taken on board at another port. Ballast water sampled from five vessels in Hong Kong Harbor contained 81 species in eight animal and five protist phyla (Chu et al. 1997). Extrapolating from the international extent of shipping, Carlton (1999) estimated that >7000 species are transported daily in ballast water, including serious human pathogens. Hundreds of species of fishes and invertebrates have become widely established as a result of such ballast water introductions, including such well-known pests as the zebra mussel, Dreissena polymorpha, and the predatory cladoceran, Bythotrephes cederstroemi, in the Laurentian Great Lakes.
These and other invertebrates can drastically alter the food resource base for fishes via competition for or elimination of natural prey. An American export, the ctenophore,Mnemiopsis leidyi, was introduced via ballast water into the Black and Azov seas of Asia, where it reached densities of 180 individuals/m3. It competed with and ate native fish larvae and has been linked to loss of a $250 million anchovy fishery (Ruiz et al. 1997; Shiganova & Bulgakova 2000). Globally, at least 32 introduced fish species in 11 families are thought to have been transported by ballast tanks; 31 species have actually been collected from ballast tanks. Gobies and blennies are the two families most commonly associated with ballast water (Wonham et al. 2000). The Yellowfin Goby, Acanthogobius flavimanus, an east Asian native, has become one of the most common benthic fishes in the San Francisco Bay–Sacramento River area. Round Goby, Neogobius melanostomus, and Tubenose Goby,

Fig. 26.9
Sentiment expressed on a bumper sticker, Burlington, Washington. Photo by G. Helfman.
Round Gobies reach densities of up to 133/m2 and are egg predators. At high densities, they could compete with native sculpin and affect benthic spawners such as Lake Sturgeon, Acipenser fulvescens, and Lake Trout, Salvelinus namaycush, thus compromising expensive rehabilitation efforts (e.g., Moyle 1991; Dubs & Corkum 1996; Chotkowski & Marsden 1999; K. M. Jones, pers. comm.).
Assessing the potential impacts of introductions
Many species introductions, including those precipitating some of the worst-case ecological scenarios, occur in developing nations where the focus is on human economic and nutritional problems. In addition to the negative ecological consequences of many introductions, traditional fishing methods are frequently displaced by introduced species, requiring new harvesting technologies or replacing local artisanal fishers with commercial or sports fishers. Although many developing nations are in desperate need of capital and of animal protein sources, simple planning measures and attention to natural distributions and local fishing techniques could often minimize results that are destructive to both the local biota and culture (e.g., CRC 2006). Whenever an introduction is being considered, be it transplantation or exotic, a protocol such as that outlined by Kohler and Courtenay (1986a, 1986b) should be followed to assess the potential biological and sociological costs and benefits of the introduction.
Related Topics