Chapter: Essential Microbiology: Protista
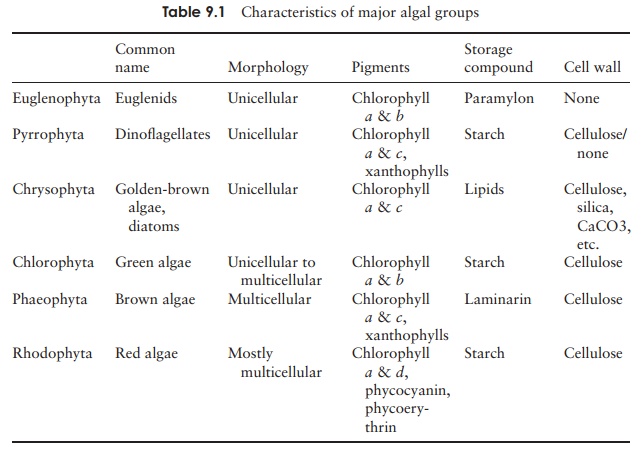
Phaeophyta - Structural characteristics of algal protists
Phaeophyta
The brown algae are multicellular, large and complex
seaweeds, which dominate rocky shores in temperate and polar regions. Apart
from one or two freshwater types, they are all marine. The presence of
fucoxanthin masks the presence of chlorophylls a and c. (In this context
it must be stated here that not all ‘brown’ seaweeds look brown, norindeed do
all the ‘red’ ones look red). Unlike the higher plants and green algae, which
use starch as a food reserve, the phaeophytes use an unusual polysaccharide
calledlaminarin (β-1,3-glucan).
The level of tissue organisation in the brown algae
is greatly in advance of any of the types we’ve discussed so far. The simplest thallus of a brown alga resembles the
most complex found in the greens.
The phaeophytes also represent an advance in terms of
sexual reproduction; here oogamy is the usual state of affairs and alternation
of generations has developed to such an extent that diploid and haploid stages
frequentlyassume separate morphological forms. Again, we shall use two examples
to illustrate life cycle diversity in the brown algae.
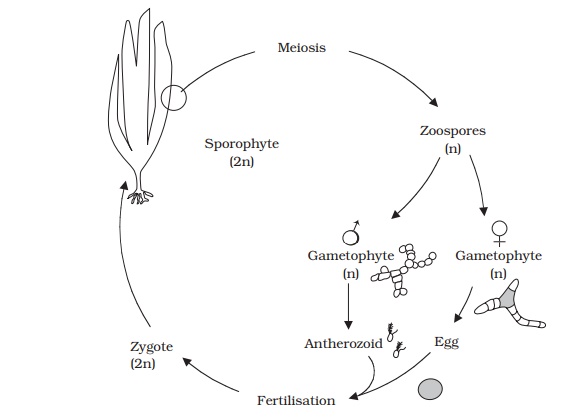
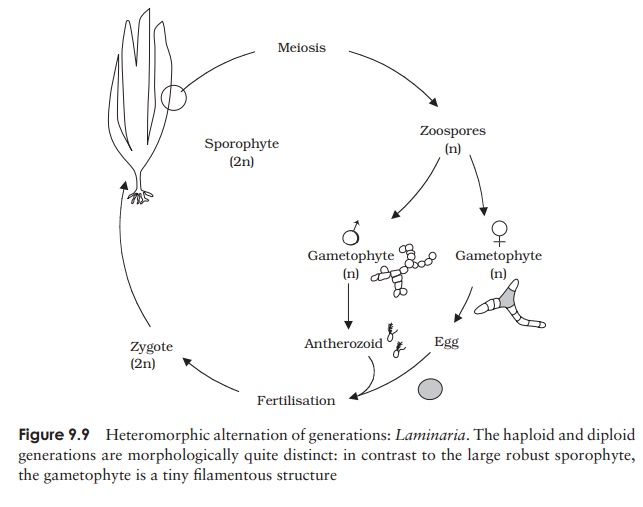
Laminaria is one of the
kelps, the largest group of brown algae. It grows attachedto underwater rocks
or other objects by means of holdfasts, root-like structures which anchor the
plant. The thallus is further subdivided into a stalk-like stipe and a broader,
blade-like lamina. Reproduction in Laminaria
involves sporophyte and gametophyte plants that are morphologically quite
distinct; (heteromorphic alternation of genera-tions). Reproductive areas
called sori develop on the blade of
the diploid sporophyte at certain times of year (Figure 9.9). These consist of
many sporangia, interspersed with thick protective hairs called paraphyses. As
the sori develop, meiosis occurs, leading to the production of haploid
zoospores. These in turn develop into haploid filamentous gametophyte plants,
much smaller and quite different in morphology from the more highly organised
sporophyte. Indeed, in contrast to the large sporophyte the gameto-phyte is a
microscopic structure. The gametophytes are dioecious, that is the male and
female reproductive structures are borne on separate individuals. The female plant
bears a number of oogonia, each of
which produces a single egg, which escapes through a pore at the apex of the
oogonium, but remains attached in a sort of cup, formed by the surrounds of the
pore. In similar fashion the male plant bears several antheridia, each
liberating a single antherozoid; this however is motile by means of flagella
and fertilises the egg. The diploid zygote so produced grows immediately into a
new sporophyte plant.
In our second example of a phaeophyte life cycle,
there is no alternation of generations at all, the gametophyte generation
having been completely lost. The wracks are familiar seaweeds found in the
intertidal zone, and Fucus vesiculosus,
known commonly as the bladder wrack, is one of the best known (Figure 9.10). It
gets its name from the air bladders distributed on its surface, which assist
buoyancy.
The adult has reproductive structures called
receptacles, slight swellings situated at the tip of the thallus; within these
are flask-like invaginations called conceptacles
which contain the male or female gametangia, again interspersed with sterile
paraphyses. F.vesiculosus is
monoecious but some other Fucus species
are dioecious. Each antheridium
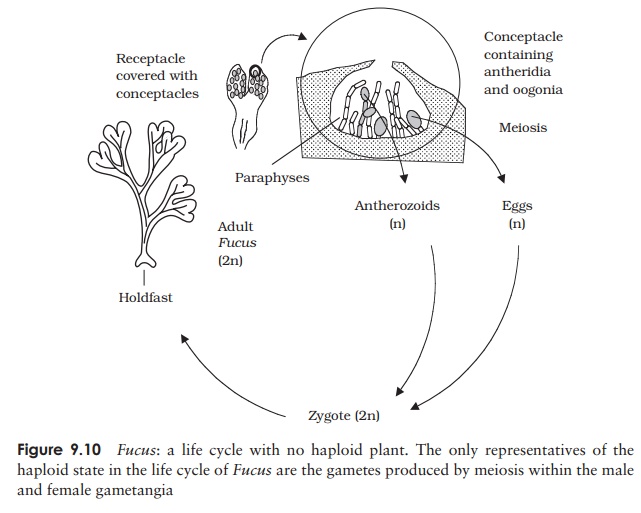
undergoes meiosis, followed by mitosis to produce 64
antherozoids or sperm, while by the same means eight eggs are produced in the
oogonium. At high tide these gametes are released into the open water.
Fertilisation results in a diploid zygote, which continues to drift quite free,
while secreting a mucilaginous covering. It eventually settles, becoming
anchored by the mucilage, and germinates into an adult individual. Here then,
we have a life cycle in which there is no gametophyte generation, and no
specialised asexual reproduction (although in certain conditions fragments may
regenerate to form adults).
Related Topics