Chapter: Essential Microbiology: Protista
Chlorophyta - Structural characteristics of algal protists
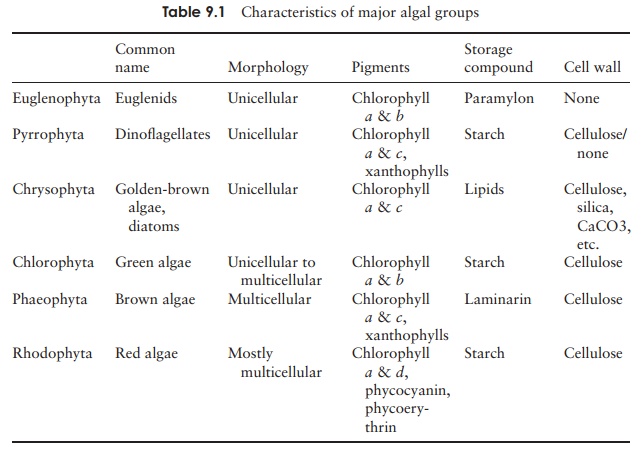
Chlorophyta
The green algae have always attracted a lot of
interest because, as a group, they share a good deal in common with the higher
plants in terms of ultrastructure, metabolism and photosynthetic pigments,
pointing to the likelihood of a common ancestor. They possess both chlorophyll a and b and certain carotenoids, store carbohydrate in the form of
starch, and generally have a rigid cell wall containing cellulose. The starch
is stored in structures called pyrenoids,
which are found within chloroplasts. There are two phylogenetically distinct
lines of green algae, the Charophyta and the Chlorophyta; the latter are much
the bigger group, but the charophytes seem to be more closely related to green
plants (see Figure 9.18).
Chlorophytes demonstrate a wide variety of body
forms, ranging from unicellular types to colonial, filamentous, membranous and
tubular forms. The vast majority of species are freshwater aquatic, but a few
marine and pseudoterrestrial representatives exist.
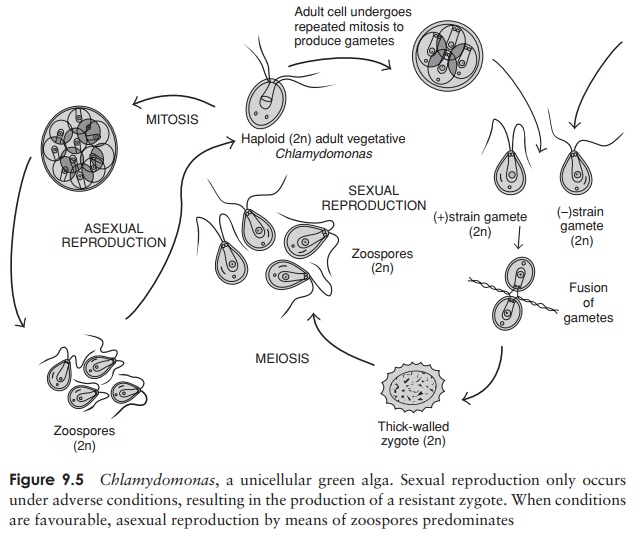
The genus usually chosen to illustrate the
unicellular condition in chlorophytes is Chlamydomonas
(Figure 9.5). This has a single chloroplast, similar in structure andshape
to that of a higher plant, and containing a pyrenoid. Situated together at the
anterior end is a pair of smooth or whiplash flagella, whose regular, ordered
contractions propel it through the water. A further structural feature found in
Chlamydomonas and other motile forms
of green algae is the stigma or eye-spot; this is made up of granules of a
carotenoid pigment and is at least partially responsible for orienting the cell
with respect to light.
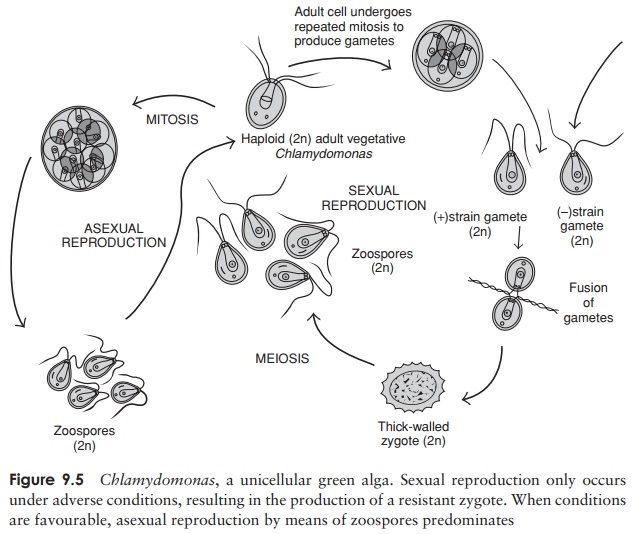
Reproduction in Chlamydomonas
and other unicellular types under favourable con-ditions of light, temperature
and nutrients, occurs asexually by the production of zoospores. A single haploid adult loses its flagella and undergoes
mitosis to produceseveral daughter cells, which then secrete cell walls and
flagella and take up an in-dependent existence of their own. This can result in
a tremendous increase in num-bers; a single cell can divide as many as eight
times in one day. Sexual reproduc-tion in Chlamydomonas,
which occurs when conditions are less favourable, differs
Any
one of the three variants of ga-mete production seen in the algae may be seen:
isogamy, anisogamy and oogamy. In all cases, two haploid gametes undergo a
fusion of both cytoplasm and nuclei to give a diploid zygote. The gametes may
simply be unmodified haploid adult cells, or they may arise through mitotic
cleavage of the adult, depending on the species. The process of isogamy, where
the two gametes are morphologically alike and cannot be differentiated
visually, only occurs in relatively lowly organisms such as Chlamydomonas. In some species we see
the beginnings of sexual differentiation – there are two mating strains,
designated + and −, and fusion
will only take place between individuals of opposite strains. The diploid
zygote, once formed, often develops into a tough-walled protective spore called
a zygospore, which tides the organism
over conditions of cold or drought. At an appropriate time the zygospore is
stimulated to recommence the life-cycle, and meiosis occurs, to produce haploid
cells, which then mature into adult individuals.
In C. braunii,
sexual reproduction is anisogamous; a + strain
produces eight mi-crogametes and a − strain produces four macrogametes. In C. coccifera, simple oogamy occurs, in
which a vegetative cell loses its flagella, rounds off and enlarges; this acts
as the female gamete or ovum, and is fertilised by male gametes formed by other
cells.
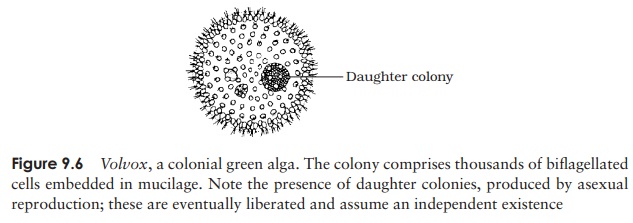
The next level of organization in the green algae is
seen in the colonial types, typified
by Volvox. These, like the
unicellular types, are motile by means of flagella, and exist as a number of
cells embedded in a jelly-like matrix (Figure 9.6). Both the number of cells and
the way they are arranged is fixed and characteristic of a particular species.
During growth, the number of cells does not increase. In simpler types, all
cells seem to be identical but in more complex forms there are distinct
anterior and posterior ends, with the stigma more prominent at the anterior,
and the posterior cells becoming larger. Reproduction can occur asexually or
sexually.
The diversity of body forms in multicellular
chlorophytes referred to earlier is matched by that of their life cycles. Two
examples are described here.
Oedogonium is a
filamentous type. When young it attaches to the substratum bya basal holdfast,
but unless it lives in flowing water the adult form is free-floating. Asexual
reproduction occurs by means of motile zoospores, which swim free for around an
hour before becoming fixed to a substratum and developing into a new filament.
In sexual reproduction, the process of sexual differentiation is carried a step
further than we have seen so far, with two separate filaments producing gametes
from specialised cells called gametangia
(Figure 9.7). These are morphologically distinct, with the male being termed an
antheridium and the female an oogonium. Gametes (morphologically
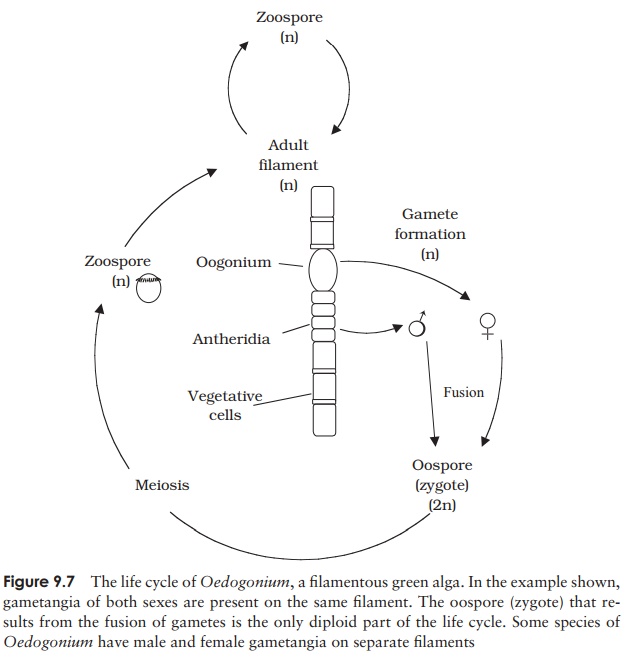
distinct: anisogamy) fuse to form a resistant zygote
or oospore, which, when conditions
are favourable, undergoes meiosis to produce four haploid zoospores, each of
which can germinate into a young haploid filament. The oospore is thus the only
diploid phase in the life cycle. In Oedogonium
there are species with separate male and female filaments (dioecious) as well as ones with both sexes on the same filament (monoecious).
A second main form of multicellularity in green algae
is the parenchymatous state, by which
we mean that the cells divide in more than one plane, giving the plant
thickness as well as length and width. An example of this is Ulva, the sea lettuce, a familiar sight
at the seaside in shallow water, attached to rocks or other objects. Ulva has a flat, membranous structure,
comprising two layers of cells. Reproductively it is of interest because it
features alternation of generations, a feature of all the higher green plants.
This means that both haploid and diploid mature individuals exist in the life
cycle. Gametes are released from one haploid adult and fuse with gametes
similarly released from another to form a zygote (Figure 9.8). In most species
of Ulva, the male and
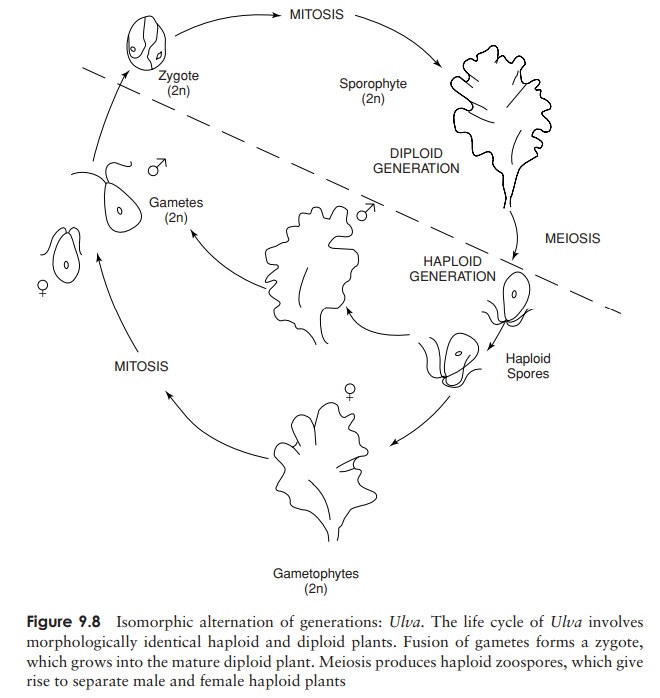
female gametes are morphologically identical
(isogamy). The zygote germinates to form a diploid plant, indistinguishable
from the plant that produced the gametes, except for its complement of
chromosomes. When the diploid plant is mature, it produces haploid zoospores by
meiosis, which settle on an appropriate substratum and develop into haploid Ulva plants. This form of alternation of
generations is called isomorphic,
because both haploid and diploid forms look alike and each assumes an equal
dominance in the life cycle. It is, however, more usual for alternation of
generations to be heteromorphic, with
the sporophyte and the gametophyte being physically dissimilar,
and with one form or other dominating.

Related Topics