Chapter: Clinical Dermatology: Disorders of pigmentation
Normal skin colour
Disorder of pigmentation
Normal skin colour
The colour of normal skin comes from a mixture of pigments. Untanned Caucasoid skin is pink, from oxyhaemoglobin in the blood within the papillary loops and superficial horizontal plexus (see Fig. 2.10). Melanin blends with this colour, e.g. after a suntan. Melanin is, of course, also responsible for the shades of brown seen in Negroid skin. Other hues are caused by the addition to these pigments of yellow from carotene, found mainly in subcutaneous fat and in the horny layer of the epidermis. There is no natural blue pigment: when blue is seen it is either because of an optical effect from normal pigment (usually melanin) in the dermis, or the presence of an abnormal pigment.
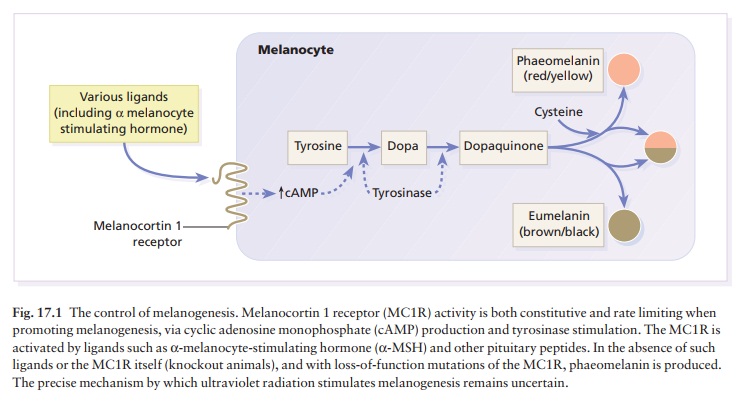
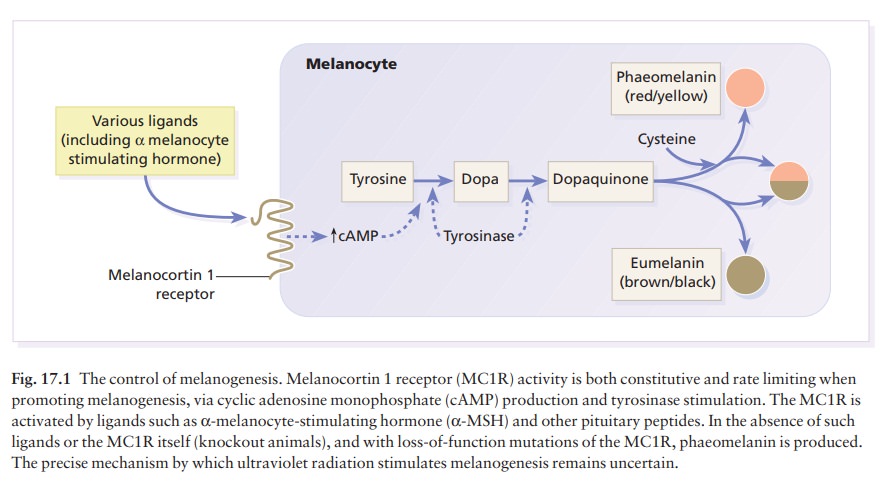
Melanogenesis
Tyrosine,
formed in the liver by hydroxylation of the essential amino acid phenylalanine
under the influence of phenylalanine hydroxylase, is the substrate for the
reactions that occur in melanocytes (Fig. 17.1). These are the only cells in
the epidermis to contain tyrosinase (dopa oxidase), the rate-limiting enzyme in
melano-genesis. This copper-dependent enzyme converts tyrosine to melanin. Dopa
is formed by the oxidation of tyrosine, and further enzymic action leads to the
formation of dopaquinone. Eumelanins, brown or black pigments, are then formed
by polymerization. Phaeomelanins and trichochromes, the pigments in red hair,
are synthesized in a similar way, except that cysteine reacts with dopaquinone
and is incorporated into the subsequent polymers. Phaeomelanins and eumelanins
may intermesh to form mixed melanin polymers.
Eumelanins
and phaeomelanins differ from neuro-melanins, the pigments found in the
substantia nigra and in cells of the chromaffin system (adrenal medulla,
sympathetic ganglia, etc.). The latter are derived from tyrosine using a
different enzyme, tyrosine hydroxy-lase, which is not found in melanocytes.
Melanin
is made within melanosomes, tiny part-icles measuring about 0.1 × 0.7 µm, shaped either like American footballs
(eumelanosomes, containing eume-lanin) or British soccer balls
(phaeomelanosomes, containing phaeomelanin). Eventually, fully melanized
melanosomes pass into the dendritic processes of the melanocyte to be injected
into neighbouring keratin-ocytes. Once there, the melanosomes are engulfed in
lysosomal packages (melanosome complexes) and distributed throughout the
cytoplasm.
Negroids
are not black because they have more melanocytes than Caucasoids, but because
their melanocytes produce more and larger melanosomes, which are broken down
less rapidly in the melano-some complexes. Melanins protect against ultraviolet
radiation (UVR) damage by absorbing and scattering the rays, and by scavenging
free radicals.
The control of melanogenesis
Melanogenesis
can be increased by several stimuli, the most important of which is UVR.
Tanning involves two distinct reactions.
1 An
immediate reaction occurs within 5 min ofexposure to long-wave ultraviolet
(UVA: 320–400 nm) and may be because of the photo-oxidation of preformed
melanin. This pigment-darkening reac-tion, which lasts about 15 min, is
responsible for the well-known phenomenon of a ‘false tan’.
2
The production of new pigment is delayed for some 24 h
after exposure to medium wave ultraviolet (UVB: 290–320 nm) and UVA. It depends
on the pro-liferation of melanocytes, an increase in tyrosinase activity and
melanosome production, and an increased transfer of new melanosomes to their
surrounding keratinocytes.
A
neat control mechanism involving glutathione has been postulated. Reduced
glutathione in the epi-dermis, produced by the action of glutathione reduc-tase
on glutathione, inhibits tyrosinase. UVR and some inflammatory skin conditions
may induce pig-mentation by oxidizing glutathione and so blocking its
inhibition of melanogenesis.
Melanocytes
are also influenced by melanocyte-stimulating hormone (MSH; peptides from the
pitu-itary and other areas of the brain) (Fig. 17.1). Their
melanocyte-stimulating activity is caused by a com-mon heptapeptide sequence,
cleaved from the pre-cursor protein, pro-opiomelanocortin, as a result of two
proconvertase enzymes. However, MSH peptides may play little part in the
physiological control of pig-mentation. Hypophysectomy will not cause a black
skin to lighten and only large doses of adrenocorti-cotrophic hormone (ACTH),
in pathological states , will increase skin pigmentation. It is now known that
pro-opiomelanocortin and MSH peptides are also produced by both keratinocytes and
melanocytes in the skin; so MSH may have a paracrine or autocrine function. In
the skin, α-MSH also
acts as an anti-inflammatory agent by antagonizing the effects of interleukin 1
(IL-1) in inducing IL-2 receptors on lym-phocytes and in inducing pyrexia.
Animal
experiments indicate that oestrogens, progestogens and, possibly, testosterone
may, in some circumstances, stimulate melanogenesis, either directly or by
increasing the release of MSH peptides from the pituitary.
Genetics and skin pigmentation
Genetic
differences determine the pigmentation of the different races. The black person
living in Britain, and the white person living in Africa remain black and
white, respectively. None the less, there is some phe-notypic variation in skin
colour, e.g. tanning after sun exposure. Recently, red hair has been shown to
be a result of genetic variations in the amino acid sequence of the
melanocortin 1 receptor (MC1R) (Fig. 17.1).
Related Topics