Chapter: Human Nervous System and Sensory Organs : Cerebellum
Neuronal Circuits - Structure of Cerebellum
Neuronal Circuits
Afferent Fibers (A, B)
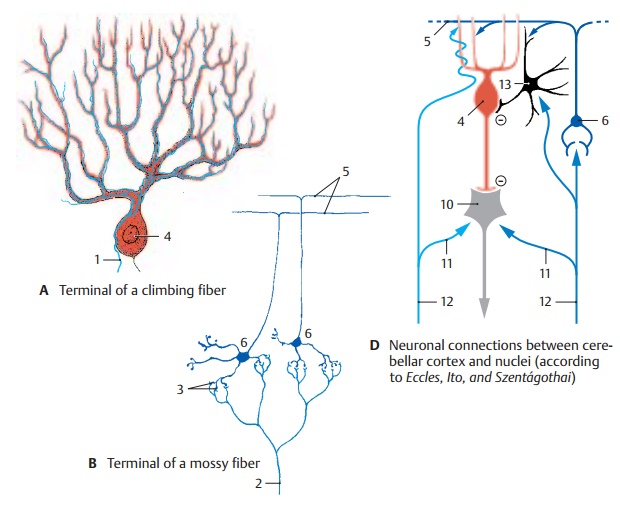
The afferent fiber systems terminate in the cerebellar cortex and give off axon collater-als to the cerebellar nuclei. There are two different types of terminals: climbing fibers and mossy fibers.
The climbing fibers (AC1) terminate at the Purkinje cells by splitting up and attaching like tendrils to the ramifications of the den-dritic tree. Each climbing fiber terminates at a single Purkinje cell and via axon collaterals also at some stellate and basket cells. The climbing fibers originate from neurons of the olive and its accessory nuclei.
The mossy fibers (BC2) divide into widely divergent branches and finally give off numerous lateral branches with small rosettes of spheroid terminals. These fit into the clawlike terminals of the granule celldendrites and form synaptic complexeswith them (B3). Mossy fibers terminate the spinocerebellar and pontocerebellar tracts and also fibers from nuclei of the medulla.
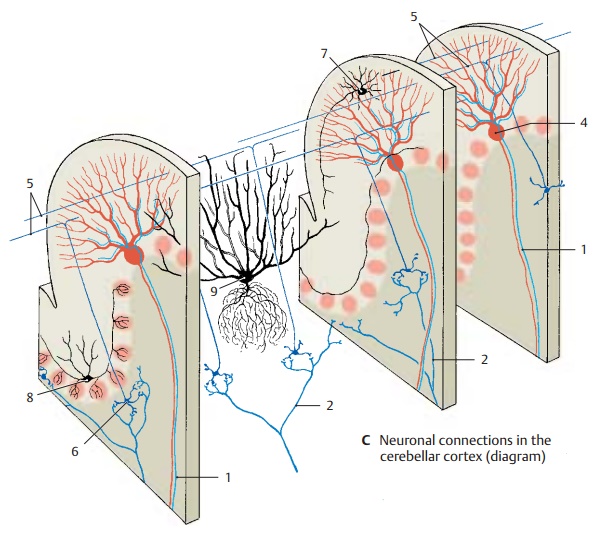
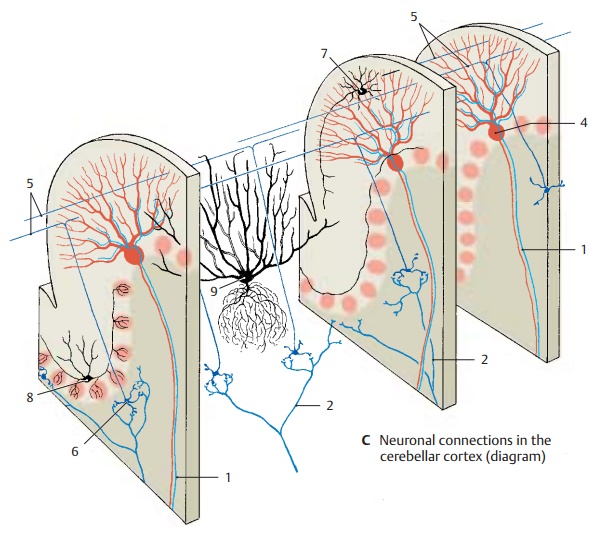
Cortex (C)
The structure of the cerebellar cortex is de-termined by the transverse orientation of the flattened dendritic trees of the Purkinje cells (ACD4) and the longitudinally extend-ing parallel fibers of the granule cells (B – D5), which form synapses with the Purkinje cell dendrites. The Purkinje cells represent the efferent elements of the cor-tex. They receive excitatory input through direct contact with climbing fibers (C1) and indirectly through mossy fibers (C2) via in-terposed granule cells (CD6). The axons of granule cells bifurcate in the molecular layer into two parallel fibers each, which measure approximately 3 mm in total length and travel through approximately 350 dendritic trees. About 200 000 parallel fibers are thought to pass through each den-dritic tree.
Stellate cells (C7), basket cells (C8), and Golgi cells (C9) are inhibitory inter-neurons which inhibit Purkinje cells. Theyare coexcited by every incoming impulse, either via synapses in the glomeruli, via synapses with parallel fibers, via synapses of Golgi cells with mossy fibers, or via axon collaterals of afferent fibers. In this way, the excitation of one row of Purkinje cells inhib-its all neighboring Purkinje cells; this sharp-ens the boundaries between active Purkinje cell clusters (contrast formation).
Functional Principle of the Cerebellum (D)
The axons of the Purkinje cells (D4) termi-nate at the neurons of the subcortical nuclei (D10) (cerebellar nuclei and vestibular nu-clei).Purkinje cells are inhibitory neurons with a high GABA content. They have a strong inhibitory effect on the neurons of the cerebellar nuclei, which continuously receive excitatory input exclusively via axon collaterals (D11) of the afferent fibers (climbing fibers and mossy fibers) (D12). However, these impulses cannot be passed on further, because the nuclei are under the inhibitory control of the Purkinje cells. Only when Purkinje cells are inhibited by inhibi-tory interneurons (D13) is their inhibiting effect lifted so that excitation is passed on to the corresponding segments of the nuclei.
Hence, the cerebellar nuclei are independentsynaptic centers, which receive and transmitimpulses and in which there is continuous excitation. Transmission is regulated by the cerebellar cortex by means of fine-tuned in-hibition and disinhibition.
Related Topics