Chapter: Human Nervous System and Sensory Organs : Cerebellum
Cerebellar Cortex - Structure of Cerebellum
Cerebellar Cortex
Overview (A)
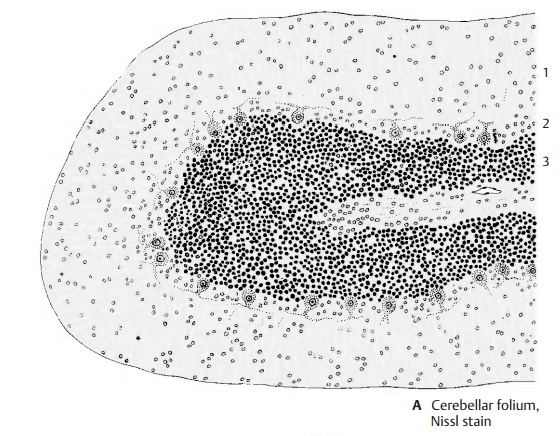
The cortex lies immediately below the sur-face and follows the course of the sulci and folia. Projection of the convoluted relief of the human cerebellum onto a plane results in an expanse of 1 m in length in the oro-caudal dimension (from the lingula to the nodulus). The cortex is regularly structured throughout all regions of the cerebellum. It consists of three layers:
The molecular layer
The Purkinje cell layer
The granular layer
The molecular layer (A1) lies beneath the surface; it contains few cells and consists mainly of unmyelinated fibers. Among its neurons we can distinguish the outer stel-late cells (lying close to the surface) and theinner basket cells. The narrow Purkinje celllayer (ganglionic layer) (A2) is formed bythe large neurons of the cerebellum, the Purkinje cells. Then follows thegranularlayer (A3). It is very rich in cells, consistingof densely packed, small neurons, the granule cells. There are also scattered largercells, the Golgi cells.
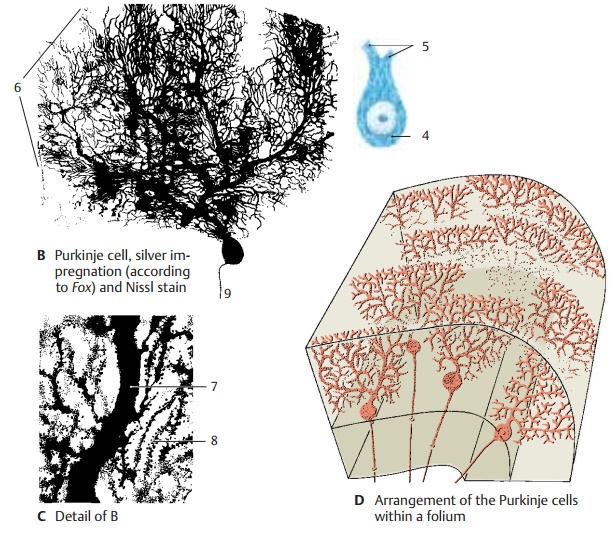
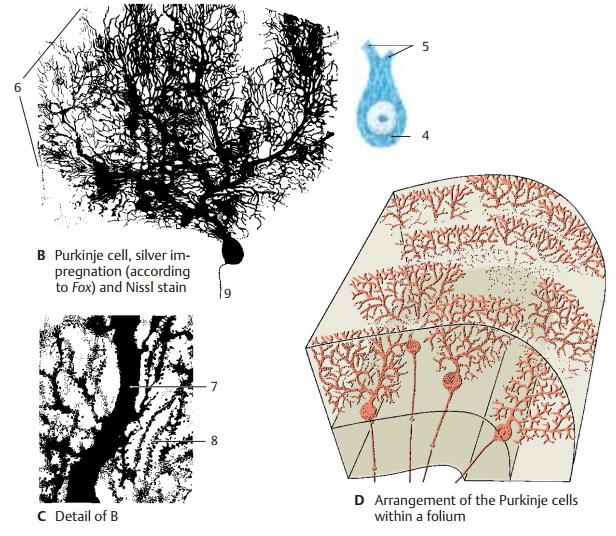
Purkinje Cells (B – D)
The Purkinje cell represents the largest and most characteristic cell of the cerebellum. The Nissl stain shows the pear-shaped cell body (B4) filled with coarse Nissl bodies. Also visible are the basal portions of two or three dendrites (B5) at the upper pole of the cell. However, the cell’s entire expanse with all its processes can only be visualized by Golgi impregnation or intracellular staining. The primary stems of the dendrites ramify into further branches, and these again into fine arborizations that form the dendritic tree (B6). The dendrites spread in a two-dimen-sional plane like the branches of an espalier tree. The Purkinje cells are arranged in a strictly geometric fashion; spaced at rela-tively regular intervals, they form a row be-tween granular and molecular layers and send their dendritic trees into the molecular layer toward the surface of the folium. Without exception, the flattened dendritic trees extend in a plane perpendicular to the longitudinal axis of the cerebellar folium (D).
The initial branches of the dendritic tree (primary and secondary dendrites) have a smooth surface (C7) and are covered withsynapses. The fine terminal branches are dotted with short spines (C8). Each Purkinje cell carries approximately 60 000 spinoussynapses. Different fiber systems terminateat the smooth and spiny sections of the cell: the climbing fibers end at the smooth sec-tion and the parallel fibers at the spiny section.
The axon (B9) departs from the base of the Purkinje cell and extends through the granular layer into the white matter. The axons of Purkinje cells terminate at neurons of the cerebellar nuclei. They give off recurrent collaterals. Purkinje cells use GABA as neurotransmitter.
Stellate Cells and Basket Cells (A, B)
In the upper half of the molecular layer lie the stellate cells. The dendrites of these small neurons run in all directions and reach ap-proximately 12 Purkinje dendritic trees. Their axons terminate either at the Purkinje cell bodies or run horizontally below the surface of the folium.
In the lower third of the molecular layer lie the slightly larger basket cells (A1). Their long axons run horizontally above the Purkinje cell bodies and give off collaterals, the terminal branches of which form net-works (baskets) around the Purkinje cell bodies. The electron-microscopic image shows that the basket cell fibers form numerous synaptic contacts (B2) with the Purkinje cell, namely, at the base of the cell body (axon hillock) and at the initial seg-ment of the axon up to where the myelin sheath begins. The rest of the Purkinje cell body is enveloped by Bergmann’s glial cells (B3). The positioning of the synapses at the axon hillock indicates the inhibitorycharacter of the basket cells.
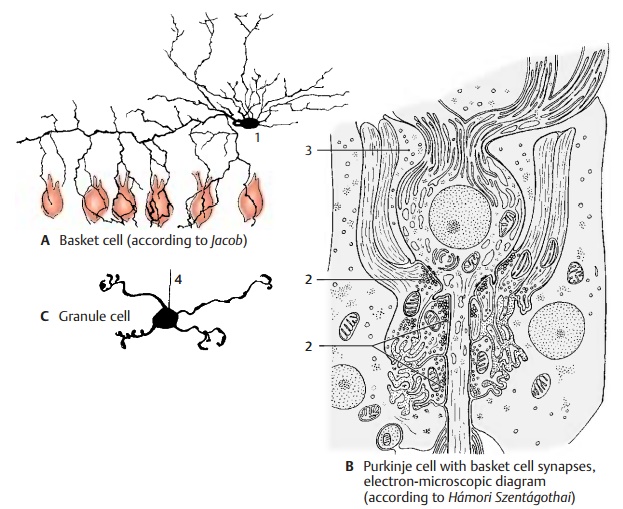
Granule Cells (C)
These small, densely packed neurons form the granular layer. At high magnification, the Golgi impregnation shows three to five short dendrites which carry clawlike thick-enings at their terminal branches. The thin axon (C4) of the granule cell ascends verti-cally through the Purkinje cell layer into the molecular layer, where it bifurcates at right angles into two parallel fibers.
Cerebellar Glomeruli.The granular layercontains small, cell-free islets (glomeruli), in which the clawlike dendritic endings of the granule cells form synaptic contacts with the axon terminals of afferent nerve fibers (mossy fibers, p. 161, B3). In addition, the short axons of the Golgi cells terminate here. The electron-microscopic image shows complex, large synapses (glomeru-lus-like synaptic complexes), which are en-veloped by glial processes
Golgi Cells (E)
These are much larger than the granule cells and are scattered throughout the granular layer, usually slightly below the Purkinje cells. Their dendritic trees, which ramify predominantly in the molecu-lar layer and extend toward the surface of the folium, are not flattened like those of the Purkinje cells but are spread in all direc-tions. The cells have short axons, which either terminate in a glomerulus or ramify into a fine, dense fiber network. The Golgi cells belong to the inhibitory interneurons.
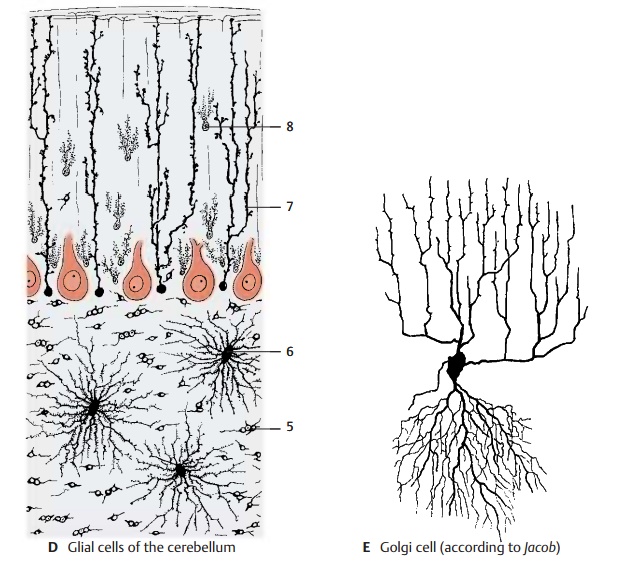
Glia (D)
Apart from the regular glial cell types, such as the oligodendrocytes (D5) and proto-plasmic astrocytes (D6) commonly found in the granular layer, there are also glial cells that are characteristic for the cerebellum: Bergmann’s glia and the penniform glia of Fañanás.
The cell bodies of the Bergmann’s cells (D7) lie between the Purkinje cells and send long supporting fibers vertically toward the sur-face, where their small end-feet form a lim-iting glial membrane against the meninges. The supporting fibers carry leaflike processes and form a dense scaffold. Berg-mann’s glia begins to proliferate where Purkinje cells go dead. The Fañanás cells (D8) have several short processes with a charac-teristic penniform structure.
Related Topics