Chapter: Medical Physiology: Contributions of the Cerebellum and Basal Ganglia to Overall Motor Control
Neuronal Circuit of the Cerebellum
Neuronal Circuit of the Cerebellum
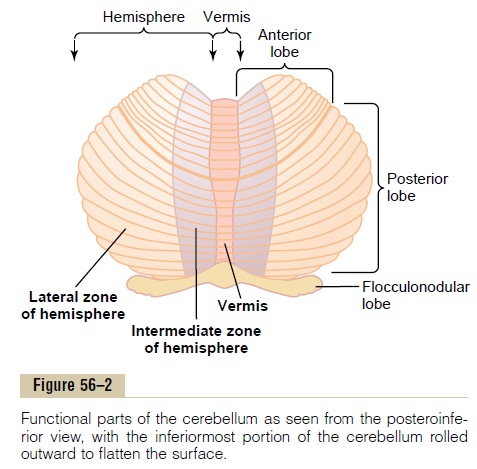
The human cerebellar cortex is actually a large folded sheet, about 17 centimeters wide by 120 centimeters long, with the folds lying crosswise, as shown in Figures 56–2 and 56–3. Each fold is called a folium. Lying deep beneath the folded mass of cerebellar cortex are deepcerebellar nuclei.
Input Pathways to the Cerebellum
Afferent Pathways from Other Parts of the Brain. The basicinput pathways to the cerebellum are shown in Figure 56–4. An extensive and important afferent pathway is the corticopontocerebellar pathway, which originates in the cerebral motor and premotor corticesand also in the cerebral somatosensory cortex. It passes by way of the pontile nuclei and pontocerebellar tracts mainly tothe lateral divisions of the cerebellar hemispheres on the opposite side of the brain from the cerebral areas.
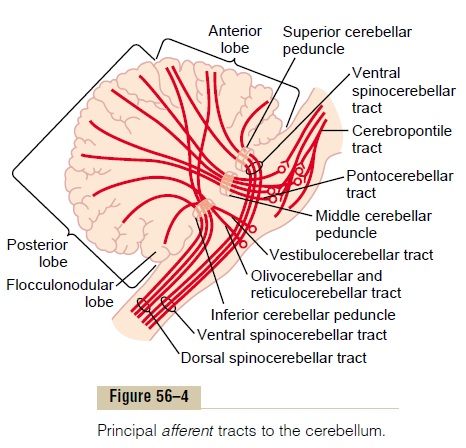
In addition, important afferent tracts originate in each side of the brain stem; they include (1) an exten-sive olivocerebellar tract, which passes from the inferiorolive to all parts of the cerebellum and is excited in theolive by fibers from the cerebral motor cortex,basalganglia, widespread areas of the reticular formation, and spinal cord; (2) vestibulocerebellar fibers, some of which originate in the vestibular apparatus itself and others from the brain stem vestibular nuclei—almost all of these terminate in theflocculonodular lobe and fastigialnucleus of the cerebellum; and (3) reticulocerebellar fibers, which originate in different portions of the brainstem reticular formation and terminate in the midline cerebellar areas (mainly in the vermis).
Afferent Pathways from the Periphery. The cerebellum alsoreceives important sensory signals directly from the peripheral parts of the body mainly through four tracts on each side, two of which are located dorsally in the cord and two ventrally. The two most important of these tracts are shown in Figure 56–5: the dorsal spinocere-bellar tract and the ventral spinocerebellar tract. Thedorsal tract enters the cerebellum through the inferior cerebellar peduncle and terminates in the vermis and intermediate zones of the cerebellum on the same side as its origin. The ventral tract enters the cerebellum through the superior cerebellar peduncle, but it termi-nates in both sides of the cerebellum.
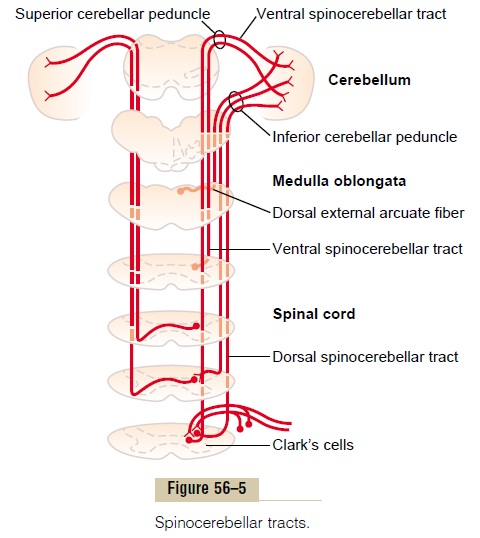
The signals transmitted in the dorsal spinocerebellar tracts come mainly from the muscle spindles and to a lesser extent from other somatic receptors throughout the body, such as Golgi tendon organs, large tactile receptors of the skin, and joint receptors. All these signals apprise the cerebellum of the momentary status of (1) muscle contraction, (2) degree of tension on the muscle tendons, (3) positions and rates of movement of the parts of the body, and (4) forces acting on the surfaces of the body.
Conversely, the ventral spinocerebellar tracts receive less information from the peripheral receptors. Instead, they are excited mainly by motor signals arriving in the anterior horns of the spinal cord from (1) the brain through the corticospinal and rubrospinal tracts and (2) the internal motor pattern generators in the cord itself. Thus, this ventral fiber pathway tells the cerebellum which motor signals have arrived at the anterior horns; this feedback is called the efference copy of the anterior horn motor drive.
The spinocerebellar pathways can transmit impulses at velocities up to 120 m/sec, which is the most rapid conduction in any pathway in the central nervous system. This extremely rapid conduction is important for instantaneous apprisal of the cerebellum of changes in peripheral muscle actions.
In addition to signals from the spinocerebellar tracts, signals are transmitted into the cerebellum from the body periphery through the spinal dorsal columns to the dorsal column nuclei of the medulla and then relayed to the cerebellum. Likewise, signals are transmitted up the spinal cord through the spinoreticular pathway to the reticular formation of the brain stem and also through the spino-olivary pathway to the inferior olivary nucleus. Then signals are relayed from both of these areas to the cerebellum. Thus, the cerebellum con-tinually collects information about the movements and positions of all parts of the body even though it is oper-ating at a subconscious level.
Output Signals from the Cerebellum
Deep Cerebellar Nuclei and the Efferent Pathways. Locateddeep in the cerebellar mass on each side are three deepcerebellar nuclei—the dentate,interposed, and fastigial. (The vestibular nuclei in the medulla also function in some respects as if they were deep cerebellar nuclei because of their direct connections with the cortex of the flocculonodular lobe.) All the deep cerebellar nuclei receive signals from two sources: (1) the cerebellar cortex and (2) the deep sensory afferent tracts to the cerebellum.
Each time an input signal arrives in the cerebellum, it divides and goes in two directions: (1) directly to one of the cerebellar deep nuclei and (2) to a corresponding area of the cerebellar cortex overlying the deep nucleus. Then, a fraction of a second later, the cerebellar cortex relays an inhibitory output signal to the deep nucleus. Thus, all input signals that enter the cerebellum even-tually end in the deep nuclei in the form of initial exci-tatory signals followed a fraction of a second later by inhibitory signals. From the deep nuclei, output signals leave the cerebellum and are distributed to other parts of the brain.
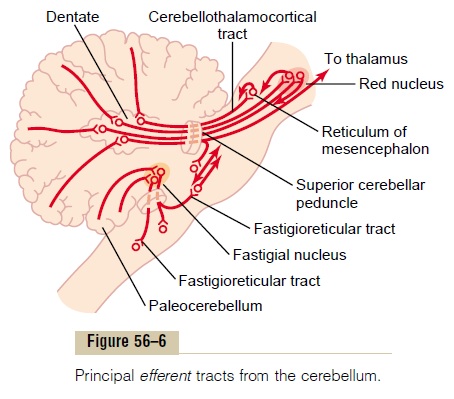
The general plan of the major efferent pathways leading out of the cerebellum is shown in Figure 56–6 and consists of the following:
1.A pathway that originates in the midline structuresof the cerebellum (the vermis) and then passesthrough the fastigial nuclei into themedullary and pontile regions of the brain stem. This circuitfunctions in close association with the equilibrium apparatus and brain stem vestibular nuclei to control equilibrium, and also in association with the reticular formation of the brain stem to control the postural attitudes of the body.
2.A pathway that originates in (1) the intermediate zone of the cerebellar hemisphere and then passes through (2) the interposed nucleus to (3) the ventrolateral and ventroanterior nuclei of the thalamus and then to (4) the cerebral cortex, to (5) several midline structures of the thalamus and then to (6) the basal ganglia and (7) the red nucleus and reticular formation of the upper portion of the brain stem. This complex circuit helps to coordinate mainly the reciprocal contractions of agonist and antagonist muscles in the peripheral portions of the limbs, especially in the hands, fingers, and thumbs.
3.A pathway that begins in the cerebellar cortex of the lateral zone of the cerebellar hemisphere and then passes to the dentate nucleus, next to the ventrolateral and ventroanterior nuclei of the thalamus, and, finally, to the cerebral cortex. This pathway plays an important role in helping coordinate sequential motor activities initiated by the cerebral cortex.
Functional Unit of the Cerebellar Cortex— The Purkinje Cell and the Deep Nuclear Cell
The cerebellum has about 30 million nearly identical functional units, one of which is shown to the left in Figure 56–7. This functional unit centers on a single, very large Purkinje cell (30 million of which are in the cerebellar cortex) and on a correspondingdeepnuclear cell.
To the top and right in Figure 56–7, the three major layers of the cerebellar cortex are shown: the molecu-lar layer, Purkinje cell layer, and granule cell layer.
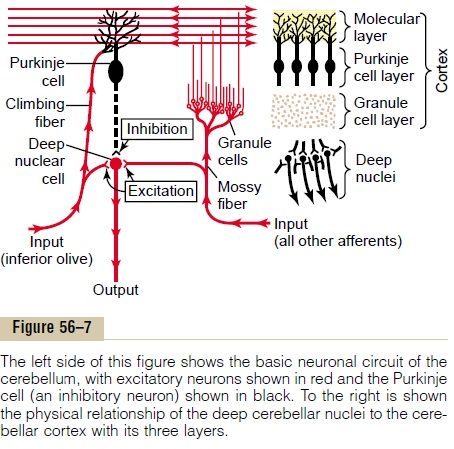
Beneath these cortical layers, in the center of the cere-bellar mass, are the deep cerebellar nuclei that send output signals to other parts of the nervous system.
Neuronal Circuit of the Functional Unit. Also shown in theleft half of Figure 56–7 is the neuronal circuit of the functional unit, which is repeated with little variation 30 million times in the cerebellum. The output from the functional unit is from a deep nuclear cell. This cell is continually under both excitatory and inhibitory influences. The excitatory influences arise from direct connections with afferent fibers that enter the cere-bellum from the brain or the periphery. The inhibitory influence arises entirely from the Purkinje cell in the cortex of the cerebellum.
The afferent inputs to the cerebellum are mainly of two types, one called the climbing fiber type and the other called the mossy fiber type.
The climbing fibers all originate from the inferiorolives of the medulla. There is one climbing fiber for about 5 to 10 Purkinje cells. After sending branches to several deep nuclear cells, the climbing fiber continues all the way to the outer layers of the cerebellar cortex, where it makes about 300 synapses with the soma and dendrites of each Purkinje cell. This climbing fiber is distinguished by the fact that a single impulse in it will always cause a single, prolonged (up to 1 second), peculiar type of action potential in each Purkinje cell with which it connects, beginning with a strong spike and followed by a trail of weakening secondary spikes. This action potential is called the complex spike.
The mossy fibers are all the other fibers that enter the cerebellum from multiple sources: from the higher brain, brain stem, and spinal cord. These fibers also send collaterals to excite the deep nuclear cells. Then they proceed to the granule cell layer of the cortex, where they too synapse with hundreds to thousands of granule cells. In turn, the granule cells send very, verysmall axons, less than 1 micrometer in diameter, up to the molecular layer on the outer surface of the cere-bellar cortex. Here the axons divide into two branches that extend 1 to 2 millimeters in each direction paral-lel to the folia. There are many millions of these par-allel nerve fibersbecause there are some 500 to 1000granule cells for every 1 Purkinje cell. It is into this molecular layer that the dendrites of the Purkinje cells project and 80,000 to 200,000 of the parallel fibers synapse with each Purkinje cell.
The mossy fiber input to the Purkinje cell is quite different from the climbing fiber input because their synaptic connections are weak, so that large numbers of mossy fibers must be stimulated simultaneously to excite the Purkinje cell. Furthermore, activation usually takes the form of a much weaker short-duration Purkinje cell action potential called a simplespike, rather than the prolonged complex actionpotential caused by climbing fiber input.
Purkinje Cells and Deep Nuclear Cells Fire Continuously Under Normal Resting Conditions. One characteristic of bothPurkinje cells and deep nuclear cells is that normally both of them fire continuously; the Purkinje cell fires at about 50 to 100 action potentials per second, and the deep nuclear cells at much higher rates. Further-more, the output activity of both these cells can be modulated upward or downward.
Balance Between Excitation and Inhibition at the Deep Cere-bellar Nuclei. Referring again to the circuit of Figure 56–7, one should note that direct stimulation of the deep nuclear cells by both the climbing and the mossy fibers excites them. By contrast, signals arriving from the Purkinje cells inhibit them. Normally, the balance between these two effects is slightly in favor of exci-tation, so that, under quiet conditions, output from the deep nuclear cell remains relatively constant at a mod-erate level of continuous stimulation.
In execution of a rapid motor movement, the initiating signal from the cerebral motor cortex or brain stem at first greatly increases deep nuclear cell excitation. Then, another few milliseconds later, feedback inhibitory signals from the Purkinje cell circuit arrive. In this way, there is first a rapid excita-tory signal sent by the deep nuclear cells into the motor output pathway to enhance the motor move-ment, but this is followed within another small fraction of a second by an inhibitory signal. This inhibitory signal resembles a “delay-line” negative feedback signal of the type that is effective in providing damping. That is, when the motor system is excited,a negative feedback signal occurs after a short delay to stop the muscle movement from overshooting its mark. Otherwise, oscillation of the movement would occur.
Other Inhibitory Cells in the Cerebellum. In addition to thedeep nuclear cells, granule cells, and Purkinje cells, two other types of neurons are located in the cerebellum: basket cells and stellate cells. These are inhibitory cellswith short axons. Both the basket cells and the stellate cells are located in the molecular layer of the cerebel-lar cortex, lying among and stimulated by the small parallel fibers. These cells in turn send their axons at right angles across the parallel fibers and cause lateralinhibition of adjacent Purkinje cells, thus sharpeningthe signal in the same manner that lateral inhibition sharpens contrast of signals in many other neuronal circuits of the nervous system.
Turn-On/Turn-Off and Turn-Off/Turn-On Output Signals from the Cerebellum The typical function of the cerebellum is to help provide rapid turn-on signals for the agonist muscles and simultaneous reciprocal turn-off signals for the antagonist muscles at the onset of a movement. Then on approaching termination of the movement, the cerebellum is mainly responsible for timing and exe-cuting the turn-off signals to the agonists and turn-on signals to the antagonists. Although the exact details are not fully known, one can speculate from the basic cerebellar circuit of Figure 56–7 how this might work, as follows.
Let us suppose that the turn-on/turn-off pattern of agonist/antagonist contraction at the onset of move-ment begins with signals from the cerebral cortex. These signals pass through noncerebellar brain stem and cord pathways directly to the agonist muscle to begin the initial contraction.
At the same time, parallel signals are sent by way of the pontile mossy fibers into the cerebellum. One branch of each mossy fiber goes directly to deep nuclear cells in the dentate or other deep cerebellar nuclei; this instantly sends an excitatory signal back into the cerebral corticospinal motor system, either by way of return signals through the thalamus to the cere-bral cortex or by way of neuronal circuitry in the brain stem, to support the muscle contraction signal that had already been begun by the cerebral cortex. As a con-sequence, the turn-on signal, after a few milliseconds, becomes even more powerful than it was at the start because it becomes the sum of both the cortical and the cerebellar signals. This is the normal effect when the cerebellum is intact, but in the absence of the cerebellum, the secondary extra supportive signal is missing. This cerebellar support makes the turn-on muscle contraction much stronger than it would be if the cerebellum did not exist.
Now, what causes the turn-off signal for the agonist muscles at the termination of the movement? Remem-ber that all mossy fibers have a second branch that transmits signals by way of the granule cells to the cerebellar cortex and eventually, by way of “parallel” fibers, to the Purkinje cells. The Purkinje cells in turn inhibit the deep nuclear cells. This pathway passesthrough some of the smallest, slowest-conducting nerve fibers in the nervous system: that is, the parallel fibers of the cerebellar cortical molecular layer, which have diameters of only a fraction of a millimeter. Also, the signals from these fibers are weak so that they require a finite period of time to build up enough exci-tation in the dendrites of the Purkinje cell to excite it. But once the Purkinje cell is excited, it in turn sends a strong inhibitory signal to the same deep nuclear cell that had originally turned on the movement. There-fore, this helps to turn off the movement after a short time.
Thus, one can see how the complete cerebellar circuit could cause a rapid turn-on agonist muscle con-traction at the beginning of a movement and yet cause also a precisely timed turn-off of the same agonist con-traction after a given time period.
Now let us speculate on the circuit for the antago-nist muscles. Most important, remember that through-out the spinal cord there are reciprocal agonist/ antagonist circuits for virtually every movement that the cord can initiate. Therefore, these circuits are part of the basis for antagonist turn-off at the onset of movement and then turn-on at termination of move-ment, mirroring whatever occurs in the agonist muscles. But we must remember, too, that the cere-bellum contains several other types of inhibitory cells besides Purkinje cells. The functions of some of these are still to be determined; they, too, could play roles in the initial inhibition of the antagonist muscles at onset of a movement and subsequent excitation at the end of a movement.
All these mechanisms are still partly speculation. They are presented here especially to illustrate ways by which the cerebellum could cause exaggerated turn-on and turn-off signals, controlling the agonist and antagonist muscles, and controlling the timing as well.
The Purkinje Cells “Learn” to Correct Motor Errors—Role of the Climbing Fibers
The degree to which the cerebellum supports onset and offset of muscle contractions, as well as timing of contractions, must be learned by the cerebellum. Typ-ically, when a person first performs a new motor act, the degree of motor enhancement by the cerebellum at the onset of contraction, the degree or inhibition at the end of contraction, and the timing of these are almost always incorrect for precise performance of the movement. But after the act has been performed many times, the individual events become progressively more precise, sometimes requiring only a few move-ments before the desired result is achieved, but at other times requiring hundreds of movements.
How do these adjustments come about? The exact answer is not known, although it is known that sensi-tivity levels of cerebellar circuits themselves progres-sively adapt during the training process. Especially, the sensitivity of the Purkinje cells to respond to the granule cell excitation becomes altered. Furthermore, this sensitivity change is brought about by signals from the climbing fibers entering the cerebellum from the inferior olivary complex.
Under resting conditions, the climbing fibers fire about once per second. But each time they do fire, they cause extreme depolarization of the entire dendritic tree of the Purkinje cell, lasting for up to a second. During this time, the Purkinje cell fires with one initial strong output spike followed by a series of diminish-ing spikes. When a person performs a new movement for the first time, feedback signals from the muscle and joint proprioceptors will usually denote to the cere-bellum how much the actual movement fails to match the intended movement.And the climbing fiber signals in some way alter long-term sensitivity of the Purkinje cells. Over a period of time, this change in sensitivity, along with other possible “learning” functions of the cerebellum, is believed to make the timing and other aspects of cerebellar control of movements approach perfection. When this has been achieved, the climbing fibers no longer need to send “error” signals to the cerebellum to cause further change.
Related Topics