Chapter: Human Neuroanatomy(Fundamental and Clinical): The Olfactory and Limbic Regions
Hippocampal Formation
Hippocampal Formation
In the human embryo, the hippocampal formation develops in relation to the medial surface of each cerebral hemisphere close to the choroid fissure of the lateral ventricle. It is at first approximately C-shaped in accordance with the outline of the body and inferior horn of the ventricle. The upper part of the formation is, however, separated from the ventricle because of the development of the corpus callosum between the two. For the same reason, this part of the formation remains underdeveloped and is represented by a thin layer of grey matter lining the upper surface of the corpus callosum. This layer is the indusium griseum.
Within the indusium griseum are embedded two bundles of longitudinally running fibres called the medial and lateral longitudinal striae (on each side of the midline). Posteriorly, the indusiumgriseum is continuous with a thin layer of grey matter related to the inferior aspect of the splenium of the corpus callosum. This grey matter is the splenial gyrusor gyrus faciolaris. The splenial gyrus runs forwards to become continuous with the dentate gyrus present in relation to the inferior horn of the lateral ventricle.
In the region of the inferior horn of the lateral ventricle, the developing hippocampus is pushed into the cavity of the ventricle because of the great development of the neighbouring neocortex. The hippocampal formation is best developed in this region and forms thehippocampus: This term includes the dentate gyrus.
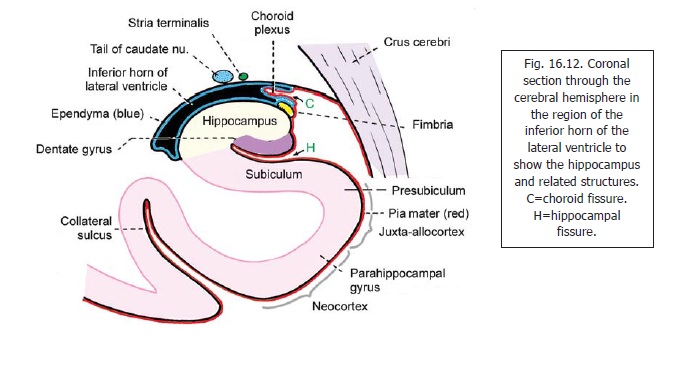
Several subdivisions of the region are described. These are illustrated in Fig. 16.12 which represents a coronal section through the inferior horn of the lateral ventricle. In this figure note that the cavity of the inferior horn is closed, on the medial side, only by apposed layers of pia mater and ependyma. A fold of pia mater (tela choroidea) projects into the ventricle and encloses a bunch of capillaries that constitute the choroid plexus. This fissure through which the tela choroidea projects into the ventricle is the choroid fissure. The cerebral cortex that lies below the choroid fissure is S-shaped in cross section. The upper and middle limbs of the ‘S’ are separated by the hippocampal fissure. The superior limb of the ‘S’ forms the hippocampus. The hippocampus consists of two parts. The part that forms the superior convex surface is calledAmmon’s horn or cornu ammonis. (This isthe hippocampus proper). The deeper part which lies below and medial to Ammon’s horn, and forms the upper wall of the hippocampal fissure, is the dentate gyrus.The middle limb of the ‘S’ connects the cornu ammonis to the parahippocampal gyrus which forms the lower limb of the ‘S’. The middle limb is an area of transition between the parahippocampal gyrus and the cornu ammonis. It is called the subiculum.
Some further subdivisions of the region are as follows. Ammon’s horn is divided into zones CA1, CA2 and CA3, CA3 being nearest the dentate gyrus, and CA1 being towards the subiculum. The subicular region is subdivided into subiculum (proper), the presubiculum, and the parasubiculum. The parasubiculum is continuous with the anterior part of the parahippocampal gyrus (entorhinal area).
The hippocampus forms a longitudinal projection that occupies the greater part of the floor of the inferior horn of the lateral ventricle. Its anterior end is expanded and notched and resembles a foot. It is therefore called the pes hippocampi.
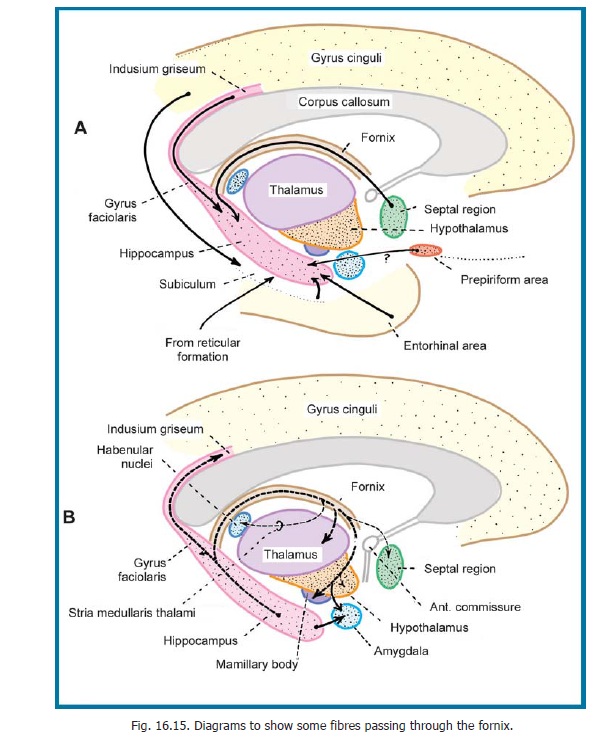
The ventricular surface of the hippocampus is covered by a layer of nerve fibres that constitute the alveus. The fibres of the alveus pass medially and collect to form a bundle of fibres, the fimbria, that projects above the medial part of the hippocampus (Fig. 16.12).
The fimbria runs backwards along the medial side of the hippocampus to become continuous with the fornix .
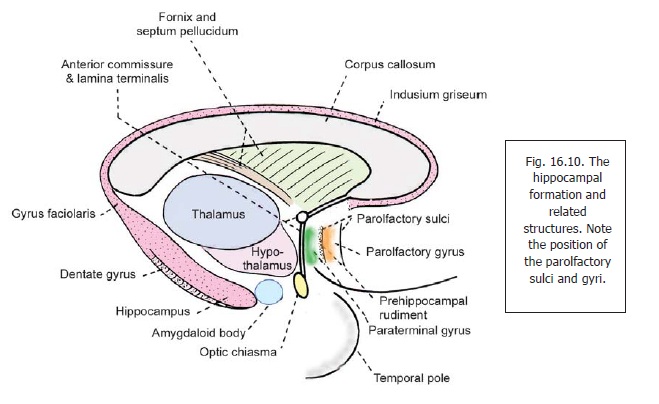
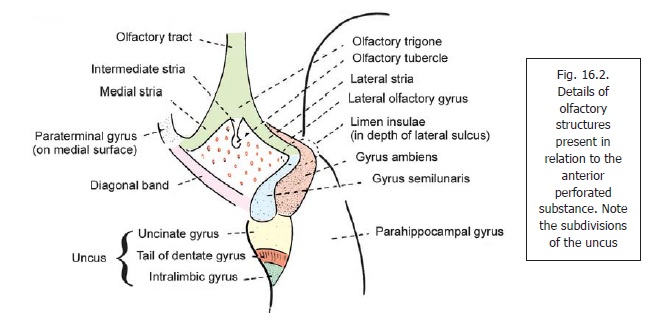
The dentate gyrus is a longitudinal strip of grey matter. Laterally, it is fused with the cornu ammonis. Its medial margin is free, and bears a series of notches that give it a dentate appearance (Fig. 16.12): hence the name dentate gyrus. When traced anteriorly the dentate gyrus runs medially across the inferior surface of the uncus. This part is called the tail of the dentate gyrus (Fig. 16.2). As stated above, the posterior end of the dentate gyrus is continuous with the splenial gyrus (gyrus faciolaris) (Fig.16.10). Because of its close relationship to the dentate gyrus the uncus is sometimes regarded as part of the hippocampal formation.
The structure of the hippocampus is different from that of the rest of the cerebral cortex. While most of the cerebral cortex is six layered the hippocampal cortex is made up of three layers only. The subiculum is a transitional zone in which a gradual reduction in the number of layers takes place. Further details of the structure of the hippocampus are beyond the scope of this section.
Connections of the hippocampus
In recent years the hippocampus has been studied in great detail and concepts regarding its connections have undergone considerable revision the main features of which are as follows.
1. Apart from its olfactory connections, the entorhinal area (area 28) is closely linked functionally tothe hippocampus, the two having numerous interconnections, and sharing their inputs and outputs .
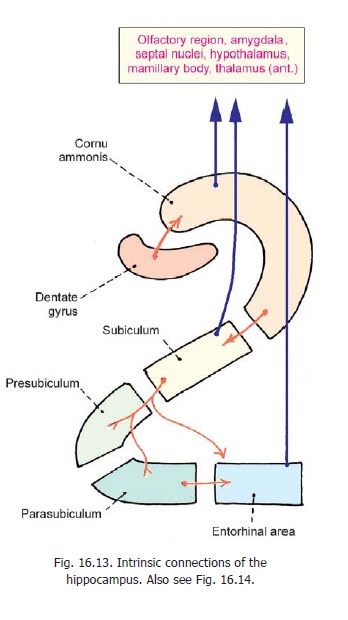
2. Intrinsic connections: The various parts of the hippocampal region are closely linked to eachother (Fig. 16.13). The dentate gyrus sends fibres to the cornu ammonis, which in turn sends fibres to the subiculum. The subiculum projects to the entorhinal area, directly or through relays in the presubiculum and parasubiculum.
3. Extrinsic connections with subcortical centres:
a. The hippocampus and entorhinal area send fibres to the amygdaloid complex and septal areas, the hypothalamus including the mamillary body, and the anterior (and some other) nuclei of the thalamus. Some fibres reach the striatum (caudate nucleus and putamen), the claustrum, and the basal nucleus of Meynert (Fig. 16.14).
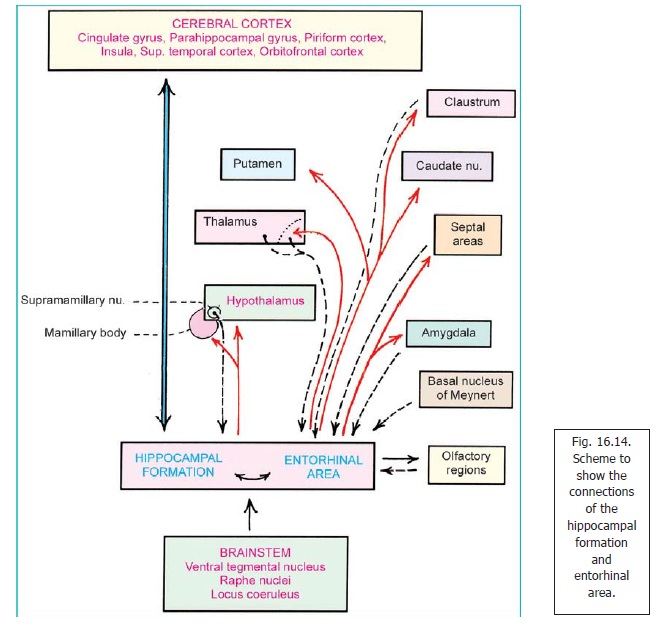
b. The hippocampus and entorhinal area receive fibres from some of the subcortical centres to which they send fibres. These include the amygdaloid complex and septal areas, the thalamus (anterior region), and the claustrum. They also receive fibres from some other regions of the thalamus and from the supramamillary nucleus (in the hypothalamus).
c. The entorhinal area receives and gives fibres to olfactory areas.
d. Fibres ascend to the hippocampus and entorhinal area from the brainstem (ventral tegmental nucleus, raphe nuclei, locus coeruleus).
4. Cortical Connections:The hippocampus and entorhinal area send fibres to and receive fibresfrom various parts of the cortex including the cingulate gyrus, the parahippocampal gyrus, the piriform cortex, the cortex of the insula, the superior temporal gyrus and the orbitofrontal cortex. In addition the region is brought into communication with many other areas of the brain through the parahippocampal gyrus.
Related Topics