Chapter: Genetics and Molecular Biology: Genes Regulating Development
General Considerations on Signaling
General Considerations on Signaling
In principle, three chemicals whose concentrations
varied in the x, y, and z directions would be sufficient to specify every
important location in a developing organism (Fig. 17.1). After determining its
position, any cell could induce or repress the genes appropriate to its
position.
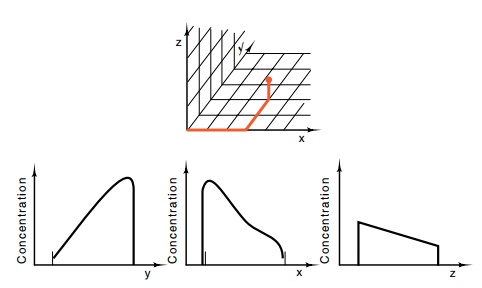
Figure 17.1 Three morphogens whose concentrations vary with position canuniquely locate a point in three dimensional space.
Generating a simple coordinate system in which the
locations of points are determined by the concentrations of three chemicals
presents several problems. The first is simply creating the gradients. How are
gradients to be built? The embryo cannot leave it to chance that an appropriate
cell will start off the process of building a gradient, perhaps by synthesizing
and secreting some compound. Therefore, either certain cells are special as a
result of their lineage and they will set up the gradients, or an external
influence directs some of the cells in the embryo to behave differently from
the rest. In either case, the embryo
Commonly, at least one external influence or
definition of an axis of the developmental coordinate system comes from the
mother. The maternal developmental signals must be special. They cannot be
intro-duced into the egg during its development if the signal is a freely
diffusing molecule. Any gradient in such a molecule would diffuse away before
fertilization and embryo development. Therefore maternal ef-fects either must
be generated during development or they must be placed in the egg in a way that
diffusion cannot alter. Another useable source of asymmetry for an embryo’s
coordinate system is the entry point of the fertilizing sperm.
To reach all cells, the chemicals used to specify
location in an embryo must be freely diffusible. If the gradients are set up in
the egg after it has been subdivided into cells, the chemicals must enter and
leave cells freely. This requires that the molecule be small. On the other
hand, position might be determined before a fertilized egg has divided into
cells. Drosophila operates this way,
and the embryo reaches about 4,000 nuclei before cell walls are synthesized.
Thus, the morphogens can be proteins.
If many different coordinate positions are to be
distinguished along one concentration gradient, precise measurements of the
morphogen concentration must be made. The standard biochemical means for measuring
concentration are simply measuring the amount of binding of a chemical to a
receptor with appropriate affinity (Fig. 17.2). This method is incapable of
discerning small differences in the concentration of a substance, but detecting
such differences would be necessary if many different developmental cues are to
be derived from one gradient. Instead of using a few gradients to determine
everything about a
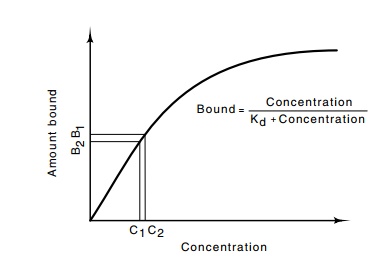
Figure
17.2 As the concentration of a
molecule increases, the amount boundto receptors increases, at first linearly
with the concentration, and then decreas-ing. Small changes in concentration
lead to small changes in the amount bound.
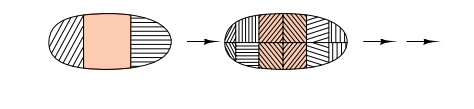
developing embryo, it is more logical that gross
patterns are first developed from simple gradients. These can be subdivided as
many times as necessary to produce as many different states as are required for
the developing embryo (Fig. 17.3).
Figure 17.3 Repeated subdivision of an embryo can generate many areas.
Not only must cells determine where they are in the
coordinate system of morphogens, but they must do so crisply. The division
lines between different tissues must be sharp. Interpenetration of one tissue
into another would generate problems. Therefore we can expect devel-oping
organisms to use special techniques to make the division lines sharp. One
simple technique is to make decisions when there are as few cells as possible.
Another is to use a small embryo, for the smaller the embryo, the steeper the
gradients, and therefore the easier it is to make decisions. Once an individual
cell has determined what tissue or body part it is to become, it can go through
multiple cell divisions to generate as much tissue as necessary. A second
general way to make sharp dividing lines in differentiating tissue is to make
the processes non-linear. First, let us examine the linear situation. Suppose
the decision between becoming tissue of type A or type B depends on the amount
of morphogen bound to a protein, and that the binding is described by the
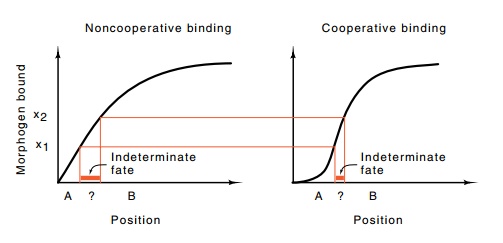
Figure
17.4 The lengths of the red boxes are
proportional to the numbers ofcells with indeterminate developmental fates.
standard Michaelis-Menten equation. Further, assume
that all cells whose receptors bind less than x1 of the morphogen become cell-type A, and all cells
whose receptors bind more than x2
of the morphogen become type B. Cells that bind an intermediate amount of
morphogen can become type A or B (Fig. 17.4). A moderate number of cells could
exist in this indeterminate position. Suppose instead, that cells use a more
complex method of measuring concentration. They could use two receptors and
respond to two gradients or respond to a single morpho-gen by requiring that
two binding events occur. Then the response of the receptor system could be
made to be the product of two Michaelis-Menton curves. This is nonlinear, and
the transition between the critical concentrations becomes much shorter.
Therefore fewer cells are sub-jected to indeterminate concentrations of
morphogen. This principle of making responses depend on multiple sensing
systems can be used to make particularly sharp responses. It is analogous to
that used by phage lambda in deciding between the lysogenic and lytic
responses.
As a
result of the general principles mentioned above, we have the following
questions concerning developmental systems. What kinds of genes and gene
products are involved? How is signaling accomplished? Are there cascades of
regulation so that subdivisions are generated within an embryo? Are multiple
gradients used in some decision proc - esses so as to make sharp boundaries?
Once a cell knows where it is, what happens next to determine its developmental
fate?
Related Topics