Chapter: Human Neuroanatomy(Fundamental and Clinical): Tracts of Spinal Cord and Brainstem
Descending Tracts ending in the Spinal Cord
Descending Tracts ending in the Spinal Cord
CORTICOSPINAL TRACTS
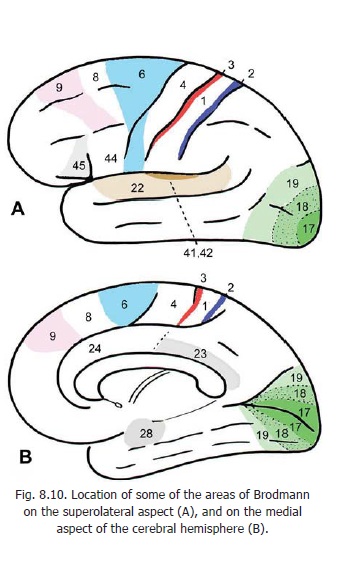
The corticospinal tracts are made up, predominantly, of axons of neurons lying in the motor area of the cerebral cortex (area 4). Some fibres also arise from the premotor area (area 6) and some from the somatosensory area (areas 3, 2, 1) (Figs. 9.3, 8.10 and 15.4). A few fibres arise in the parietal cortex (area 5). From this origin fibres pass through the corona radiata to enter the internal capsule where they lie in the posterior limb (Figs. 9.3 and 17.2A). After passing through the internal capsule the fibres enter the crus cerebri (of the midbrain): they occupy the middle two-thirds of the crus. The fibres then descend through the ventral part of the pons to enter the pyramids in the upper part of the medulla.
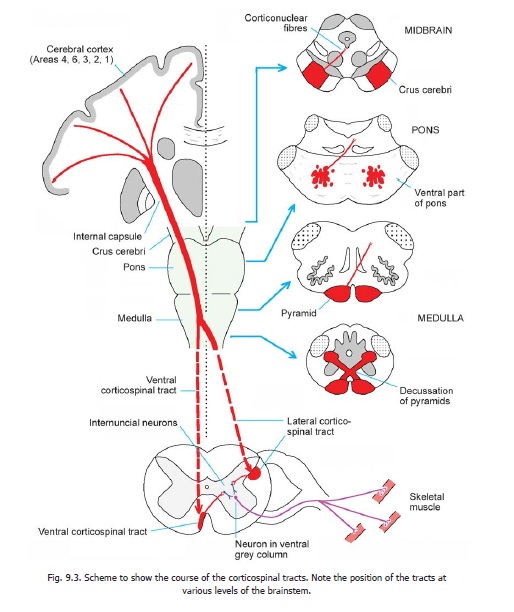
Near the lower end of the medulla about 80 percent of the fibres cross to the opposite side. (The crossing fibres of the two sides constitute the decussation of the pyramids.)
The fibres that have crossed in the medulla enter the lateral funiculus of the spinal cord and descend as the lateral corticospinal tract (Fig. 9.2). The fibres of this tract terminate in grey matter at various levels of the spinal cord. Most of them end by synapsing with internuncial neurons in the bases of the dorsal and ventral grey columns (laminae IV to VII). The internuncial neurons carry the impulses brought by fibres of the tract to ventral horn cells. Some fibres of the tract terminate directly on ventral horn cells (lamina IX, dorsolateral, central and ventrolateral groups).
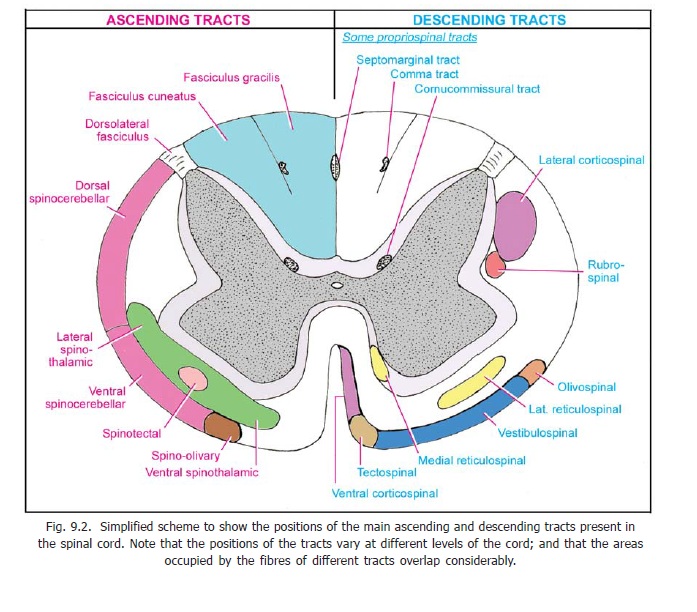
The corticospinal fibres that do not cross in the pyramidal decussation enter the anterior funiculus of the spinal cord to form theanterior corticospinal tract.
On reaching the appropriate level of the spinal cord the fibres of this tract cross the midline (through the anterior white commissure) to reach grey matter on the opposite side of the cord. Their manner of termination is similar to that of fibres of the lateral corticospinal tract. In this way the corticospinal fibres of both the lateral and anterior tracts ultimately connect the cerebral cortex of one side with ventral horn cells in the opposite half of the spinal cord.
The cerebral cortex controls voluntary movement through the corticospinal tract. Interruption of the tract anywhere in its course leads to paralysis of the muscles concerned. As the fibres are closely packed in their course through the internal capsule and brainstem small lesions here can cause widespread paralysis.
The neurons that give origin to the fibres of the corticospinal tracts are often referred to as uppermotor neurons in distinction to the ventral horn cells and their processes which constitute the lower motor neurons.Interruption of either of these neurons leads to paralysis, but the nature ofthe paralysis is distinctive in each case.
Advanced:
Having considered the basic facts about the corticospinal tracts we may now consider some additional features of interest.
1. The majority of fibres constituting the corticospinal tracts (70%) are myelinated. Most of the fibres are of small diameter (1 to 4 μm) while about 20% of fibres are of large diameter (11 to 22 μm). The latter arise from the giant pyramidal neurons (cells of Betz) lying in the motor area of the cerebral cortex.
Fibres of the corticospinal tract are unmyelinated at birth. Myelination begins in the second postnatal week and gradually extends down the axons. It is completed in the second year. The infant acquires proper motor control only after myelination of the corticospinal tracts is completed.
2. The proportion of fibres crossing in the decussation of the pyramids is variable (75 to 90%). Very rarely all corticospinal fibres may descend in the anterior corticospinal tract. Conversely, this tract may be absent. Some fibres may descend in the lateral funiculus of the same side. According to some authorities such fibres form a separate anterolateral corticospinal tract. These fibres terminate on the same side of the spinal cord.
3. The lateral corticospinal tract extends to the lowest segments of the spinal cord, but the anterior tract extends only to the midthoracic level.
4. The longest fibres of the tract (for lowest segments of the spinal cord) lie most superficially within the tract; while the shortest fibres lie most medially. Fibres of the corticospinal tracts are arranged somatotropically (This term means that within the tract the fibres meant for control of muscles in different parts of the body are arranged in a definite sequence).
5. Corticospinal fibres are usually described as ending (directly, or through internuncial neurons) in relation to alpha neurons, in contrast to some other descending tracts which are said to end in relation to gamma neurons. However, recent work shows that corticospinal fibres end in relation to both alpha and gamma neurons.
6. The influence of this tract is said to be facilitatory for flexors and inhibitory for extensors.
7. Apart from their motor functions, corticospinal fibres influence conduction in ascending tracts. This influence is mediated through axons of cells lying in the parietal lobe, which end predominately in laminae IV to VI.
8. For effects of damage to corticospinal tracts.
Rubrospinal tract
This tract is made up of axons of neurons lying in the red nucleus (which lies in the upper part of the midbrain). The fibres of the tract cross to the opposite side in the lower part of the tegmentum of the midbrain. The crossing fibres constitute the ventral tegmental decussation (Fig. 11.9). The tract descends through the pons and medulla to enter the lateral funiculus of the spinal cord (Fig. 9.2). Here the tract lies just in front of the lateral corticospinal tract. The fibres of the rubrospinal tract end by synapsing with ventral horn cells through internuncial neurons located in laminae V to VII of the spinal grey matter.
Tectospinal tract
The fibres of this tract arise from neurons in the superior colliculus (midbrain). The fibres cross to the opposite side in the upper part of the tegmentum of the midbrain. The crossing fibres form the dorsaltegmental decussation (Fig. 11.9). The tract descends through the pons and medulla into theanterior funiculus of the spinal cord. The fibres terminate by synapsing with ventral horn cells in cervical segments of the cord, through internuncial neurons located in laminae VI to VIII of the spinal grey matter.
Vestibulospinal tracts
There are two vestibulospinal tracts, lateral and medial.
Lateral vestibulospinal tract
The neurons of origin of the lateral vestibulospinal tract lie in the lateral vestibular nucleus. This tract is uncrossed and lies in the anterior funiculus of the spinal cord (Fig. 9.2) (shifting medially as it descends). Its fibres end in relation to neurons in the ventral grey column (laminae VII and VIII). This tract is an important efferent path for equilibrium.
Medial vestibulospinal tract
The medial vestibulospinal tract arises mainly from the medial vestibular nucleus (with some fibres from the inferior and lateral nuclei). The tract descends through the anterior funiculus (within the sulcomarginal fasciculus). The fibres are partly crossed and partly uncrossed. They end in cervical segments of the cord in laminae VII and VIII.
The lateral vestibulospinal tract is facilitatory to motor neurons supplying extensor muscles (of the neck, back and limbs); and is inhibitory to flexor muscles. The medial tract is inhibitory to muscles of the neck and back.
Olivospinal tract
This tract is generally described as arising from the inferior olivary nucleus (medulla) and terminating in relation to ventral horn cells of the spinal cord. However, recent research suggests that fibres do not descend from the inferior olive to the spinal cord.
Reticulospinal tracts
The reticular formation is connected to spinal grey matter through the medial and lateral reticulospinal tracts.
Medial reticulospinal tract
Fibres arise from the medial part of the reticular formation of both the pons and the medulla (mainly from the nucleus gigantocellularis reticularis of the medulla, and the oral and caudal reticular nuclei of the pons). The fibres which are crossed and uncrossed descend in the anterior funiculus (near the anterior median fissure). The fibres reach all levels of the spinal cord. They end directly, or through interneurons, on alpha and gamma motor neurons. The tract is facilitatory to muscles of the trunk and limbs, but some fibres are inhibitory to neck muscles. The tract is concerned with postural adjustments of the head, trunk and limbs.
Lateral reticulospinal tract
This tract is constituted by fibres arising in the ventrolateral part of the reticular formation of the pons (caudal and oral pontine reticular nuclei). The fibres cross to the opposite side in the medulla and run down in the lateral funiculus.
Reticulospinal fibres terminate widely in spinal grey matter, but the exact laminae in which they end are controversial. They possibly terminate in all laminae other than II and III, with the majority ending in laminae VI to VIII. Some fibres reach the lateral cervical nucleus. Apart from control of motor function, the reticulospinal tracts may influence transmission of pain through ascending tracts.
Descending Autonomic Fibres
Hypothalamospinal fibres begin (mainly) in the paraventricular nucleus of the hypothalamus,and descend uncrossed in the dorsolateral funiculus. The axons end in relation to sympathetic and parasympathetic preganglionic neurons located in the intermediolateral grey column (and other regions) of spinal grey matter. The neuromediators demonstrated in these fibres include dopamine, oxytocin and vasopressin.
Some fibres (noradrenergic) descend into the cord from the locus coeruleus. These fibres are widely distributed in spinal grey matter, but the greatest number end in relation to preganglionic parasympathetic neurons in sacral segments of the spinal cord. Some adrenergic fibres also descend from the medulla to the intermediolateral grey column.
Some groups of neurons located in the brainstem give off serotoninergic fibres that descend into the spinal cord as two bundles, lateral and ventral. The lateral fibres end in laminae I, II and V and probably influence pain perception. The ventral fibres reach laminae VIII and IX. They are facilitatory to motor neurons.
Other descending tracts
In addition to the better known descending tracts described above, fibres may also descend to the spinal cord from the interstitial nucleus (located near the third ventricle), the tegmentum of the midbrain, and from the nucleus of the solitary tract.
Significance of Descending tracts
The various descending tracts described above, that end in relation to ventral horn cells, influence their activity, and thereby have an effect on contraction and tone of skeletal muscle. Although a small number of the fibres of these tracts may synapse directly with ventral horn cells, most of them influence these cells through intervening internuncial neurons. This influence is exerted both on alpha neurons and gamma neurons. We have already seen that gamma neurons indirectly influence the activity of alpha neurons via muscle spindles. Hence, all these influences ultimately reach the alpha neurons. Such influences may be either facilitatory or inhibitory. The corticospinal and rubrospinal tracts are described as being facilitatory to flexors and inhibitory to extensors, while the vestibulospinal tract is said to have the opposite effect. The medial reticulospinal tract is generally regarded as facilitatory and the lateral tract as inhibitory. However, motor control is far more complex than such simple statements may suggest.
The corticospinal tracts are often referred to as pyramidal tracts. Traditionally all other descending tracts projecting on motor neurons have been collectively referred to as extrapyramidal tracts. It has often been presumed that the pyramidal and extrapyramidal tracts act in opposition to each other. It has also been said that the pyramidal fibres end in relation to alpha neurons, and extrapyramidal fibres in relation to gamma neurons. However, it is now recognised that such a distinction is artificial and of little significance either physiological or clinical.
We have seen that autonomic nerve fibres arise from neurons located in the general visceral efferent nuclei of the brainstem, and from the intermediolateral region of the spinal grey matter. These regions are under control from higher centres through descending autonomic pathways described above.
In addition to their influence on motor activity, it has been relatively recently recognised that descending tracts may influence the transmission of afferent impulses through ascending tracts.
Related Topics