Chapter: Medical Physiology: Contraction and Excitation of Smooth Muscle
Contractile Mechanism in Smooth Muscle
Contractile Mechanism in Smooth Muscle
Chemical Basis for Smooth Muscle Contraction
Smooth muscle contains both actin and myosin fila-ments, having chemical characteristics similar to thoseof the actin and myosin filaments in skeletal muscle. It does not contain the normal troponin complex that is required in the control of skeletal muscle con-traction, so the mechanism for control of contraction is different.
Chemical studies have shown that actin and myosin filaments derived from smooth muscle interact with each other in much the same way that they do in skele-tal muscle. Further, the contractile process is activated by calcium ions, and adenosine triphosphate (ATP) is degraded to adenosine diphosphate (ADP) to provide the energy for contraction.
There are, however, major differences between the physical organization of smooth muscle and that of skeletal muscle, as well as differences in excitation-contraction coupling, control of the contractile process by calcium ions, duration of contraction, and amount of energy required for contraction.
Physical Basis for Smooth Muscle Contraction
Smooth muscle does not have the same striated arrangement of actin and myosin filaments as is found in skeletal muscle. Instead, electron micrographic tech-niques suggest the physical organization exhibited in Figure 8–2. This figure shows large numbers of actin filaments attached to so-called dense bodies. Some of these bodies are attached to the cell membrane. Others are dispersed inside the cell. Some of the mem-brane dense bodies of adjacent cells are bonded together by intercellular protein bridges. It is mainly through these bonds that the force of contraction is transmitted from one cell to the next.
Interspersed among the actin filaments in the muscle fiber are myosin filaments. These have a diam-eter more than twice that of the actin filaments. In electron micrographs, one usually finds 5 to 10 times as many actin filaments as myosin filaments.
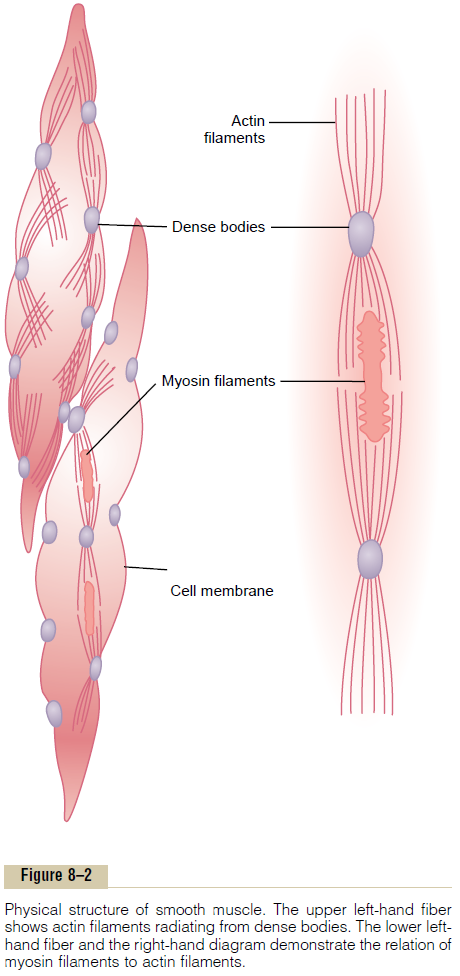
To the right in Figure 8–2 is a postulated structure of an individual contractile unit within a smooth muscle cell, showing large numbers of actin filaments radiating from two dense bodies; the ends of these filaments overlap a myosin filament located midway between the dense bodies. This contractile unit is similar to the contractile unit of skeletal muscle, but without the regularity of the skeletal muscle structure; in fact, the dense bodies of smooth muscle serve the same role as the Z discs in skeletal muscle.
There is another difference: Most of the myosin fila-ments have what are called “sidepolar” cross-bridges arranged so that the bridges on one side hinge in one direction and those on the other side hinge in the opposite direction. This allows the myosin to pull an actin filament in one direction on one side while simul-taneously pulling another actin filament in the oppo-site direction on the other side. The value of this organization is that it allows smooth muscle cells to contract as much as 80 per cent of their length instead of being limited to less than 30 per cent, as occurs in skeletal muscle.
Comparison of Smooth Muscle Contraction and Skeletal Muscle Contraction
Although most skeletal muscles contract and relax rapidly, most smooth muscle contraction is prolonged tonic contraction, sometimes lasting hours or even days.Therefore, it is to be expected that both the physi-cal and the chemical characteristics of smooth muscle versus skeletal muscle contraction would differ. Following are some of the differences.
Slow Cycling of the Myosin Cross-Bridges. The rapidityof cycling of the myosin cross-bridges in smooth muscle—that is, their attachment to actin, then release from the actin, and reattachment for the next cycle— is much, much slower in smooth muscle than in skele-tal muscle; in fact, the frequency is as little as 1/10 to 1/300 that in skeletal muscle. Yet the fraction of time that the cross-bridges remain attached to the actin fila-ments, which is a major factor that determines the force of contraction, is believed to be greatly increased in smooth muscle. A possible reason for the slow cycling is that the cross-bridge heads have far less ATPase activity than in skeletal muscle, so that degra-dation of the ATP that energizes the movements of the cross-bridge heads is greatly reduced, with corresponding slowing of the rate of cycling.
Energy Required to Sustain Smooth Muscle Contraction. Only1/10 to 1/300 as much energy is required to sustain the same tension of contraction in smooth muscle as in skeletal muscle. This, too, is believed to result from the slow attachment and detachment cycling of the cross-bridges and because only one molecule of ATP is required for each cycle, regardless of its duration.
This sparsity of energy utilization by smooth muscle is exceedingly important to the overall energy economy of the body, because organs such as the intes-tines, urinary bladder, gallbladder, and other viscera often maintain tonic muscle contraction almost indefinitely.
Slowness of Onset of Contraction and Relaxation of the Total Smooth Muscle Tissue. A typical smooth muscle tissuebegins to contract 50 to 100 milliseconds after it is excited, reaches full contraction about 0.5 second later, and then declines in contractile force in another 1 to 2 seconds, giving a total contraction time of 1 to 3 seconds. This is about 30 times as long as a single contraction of an average skeletal muscle fiber. But because there are so many types of smooth muscle, contraction of some types can be as short as 0.2 second or as long as 30 seconds.
The slow onset of contraction of smooth muscle, as well as its prolonged contraction, is caused by the slowness of attachment and detachment of the cross-bridges with the actin filaments. In addition, the initiation of contraction in response to calcium ions is much slower than in skeletal muscle, as discussed later.
Force of Muscle Contraction. Despite the relatively fewmyosin filaments in smooth muscle, and despite the slow cycling time of the cross-bridges, the maximum force of contraction of smooth muscle is often greater than that of skeletal muscle—as great as 4 to 6 kg/cm2 cross-sectional area for smooth muscle, in comparison with 3 to 4 kilograms for skeletal muscle. This great force of smooth muscle contraction results from the prolonged period of attachment of the myosin cross-bridges to the actin filaments.
“Latch” Mechanism for Prolonged Holding of Contractions of Smooth Muscle. Once smooth muscle has developed fullcontraction, the amount of continuing excitation usually can be reduced to far less than the initial level, yet the muscle maintains its full force of con-traction. Further, the energy consumed to maintain contraction is often minuscule, sometimes as little as 1/300 the energy required for comparable sustained skeletal muscle contraction. This is called the “latch” mechanism.
The importance of the latch mechanism is that it can maintain prolonged tonic contraction in smooth muscle for hours with little use of energy. Little con-tinued excitatory signal is required from nerve fibers or hormonal sources.
Stress-Relaxation of Smooth Muscle. Another impor-tant characteristic of smooth muscle, especially the vis-ceral unitary type of smooth muscle of many hollow organs, is its ability to return to nearly its original force of contraction seconds or minutes after it hasbeen elongated or shortened. For example, a sudden increase in fluid volume in the urinary bladder, thus stretching the smooth muscle in the bladder wall, causes an immediate large increase in pressure in the bladder. However, during the next 15 seconds to a minute or so, despite continued stretch of the bladder wall, the pressure returns almost exactly back to the original level. Then, when the volume is increased by another step, the same effect occurs again.
Conversely, when the volume is suddenly decreased, the pressure falls very low at first but then rises back in another few seconds or minutes to or near to the original level. These phenomena are called stress-relaxation and reverse stress-relaxation. Their impor-tance is that, except for short periods of time, they allow a hollow organ to maintain about the same amount of pressure inside its lumen despite long-term, large changes in volume.
Related Topics