Chapter: Essential Clinical Immunology: Immunological Aspects of Infection
Bacterial Infection
BACTERIAL INFECTION
The immune system responds to bacterial infections
in two major ways. First, it may respond to soluble products of the cell such
as toxins or released structural antigens like LPS of a given gram-negative
bacterial cell. Most bacterial antigens are T-cell depen-dent and require
helper T cells for initia-tion of the immune response. Yet certain cell
antigens, such as the pneumococcal polysaccharides, are T-cell independent.
They are large-molecular-weight mol-ecules, and in children, antibody response
to these antigens may take four to six years. Thus, younger children are
susceptible to these infections.
An interesting sidelight to protection against
these infections can be seen in breast-fed infants who are less susceptible to
infection than non-breast-fed babies. It now appears that it is not
polysaccharides or antibodies that are responsible for this protection but
rather a multimeric form of lactoalbumin (present in high concentra-tions in
human breast milk (see “Suggested Reading”). A broader approach to protec-tion
has been the production of pneumo-coccal polysaccharide vaccines specially
designed to induce antibody induction in the young child.
In the following discussion, strepto-cocci,
particularly S. pyogenes, are used as
the example of a bacterial infection, but many other organisms produce a
similar response. Streptococcal antigens include specific toxins such as
streptolysins O and S that lyse blood and tissue cells and pyro-genic
exotoxins, which act as superanti-gens to overstimulate the host responses.
There are also specific enzymes such as hyaluronidase and streptokinase, which
help promote the spread of infecting strep-tococcus. Perhaps most important is
the M protein (Figure 4.2), a cell surface antigen of the group A streptococcus
that allows the bacteria to evade immune defenses (especially neutrophils and
complement). One way in which M protein functions is to bind host factor H,
which prevents comple-ment C3 from depositing on the streptococ-cal surface.
Since efficient phagocytosis by neutrophils requires interaction with its C3
receptor, factor H prevents this interaction.
Antibodies to streptococcal antigens other than M
protein are slow to appear, and most likely do not play a role in lim-iting the
infection. However, antibodies to streptolysin O and deoxyribonuclease B have
become important clinical tools to determine whether a given individual has had
a recent streptococcal infection. This is partially true if a blood sample
drawn
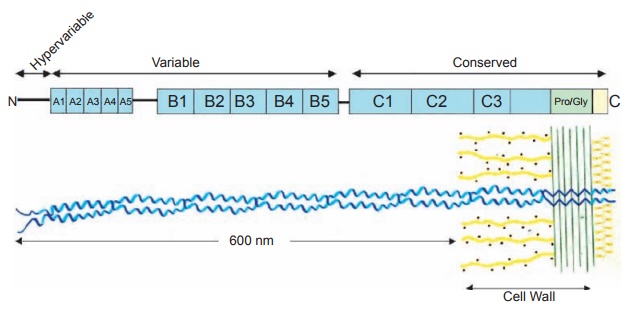
Figure 4.2 Coiled-coil
structure of the M protein. The streptococcal M protein is a coiled-coil molecule that
extends about 600 nm from the bacterial cell surface. The C-terminal region is
embedded within the cell wall and the C-terminus is located in the cytoplasmic
membrane in the nascent molecule. The coding region for the M protein is
distributed in repeat blocks designated A–C in which the C-repeat region is
conserved among M-protein serotypes, and the A and B repeats are variable among
these serotypes. The N-terminus is the hypervariable, type-specific region for
the M proteins. Pro/Gly designates the region of the M protein that is rich in
proline and glycine
at the onset and one drawn 10 to 14 days later show
a marked rise in the titer. Con-trary to the dogma, both skin infection and
pharyngeal infection with group A strepto-cocci can stimulate the production of
both antibodies.
Some bacterial antigens such as endo-toxins can be
powerful stimulators of the immune response and lead to polyclonal activation
of B lymphocytes. This rise in immunoglobulin levels is believed to be
nonspecific since only a small portion of the total immunoglobulin level is
directed to the endotoxin.
Related Topics