Chapter: Human Nervous System and Sensory Organs : Autonomic Nervous System
Autonomic Periphery of Autonomic Nervous System
Autonomic Periphery
Efferent Fibers
The preganglionic fibers are myelinated, while the postganglionic fibers are unmyeli-nated. The unmyelinated fibers are sur-rounded by the cytoplasm of Schwann cells, and a single Schwann cell envelopes several axons.
Afferent Fibers
The viscerosensory fibers are myelinated. They are regarded as being neither sympa-thetic nor parasympathetic in nature. In general, they accompany the sympathetic nerves and enter the spinal cord through the posterior roots. The fibers from the heart run through the upper thoracic roots, those from the stomach, liver, and gallblad-der run through the middle thoracic roots, and those from the colon and appendix run through the lower thoracic roots. The re-spective dermatomes of these roots corre-spond roughly to the various Head’s zones.
Intramural Plexus (A – E)
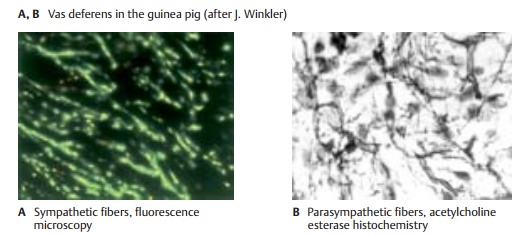
The autonomic nerves enter the internal or-gans together with the vessels and form a fine network of noradrenergic (A) or cholin-ergic (B) fibers (enteric plexus). The fibers terminate on smooth-muscle cells and on glands. Vascular muscles influence the function of many organs (regulation of blood flow by contraction or dilatation of vessels). It is not known whether parenchy-mal organs such as liver or kidneys contain secretory fibers.
The digestive tract is supplied by two differ-ent plexuses, namely, the submucous plexus (Meissner’s plexus) and the myenteric plexus (Auerbach’s plexus). The submucous plexus(C1, D) forms a three-dimensional network throughout the entire submucosa. It is an ir-regular meshwork of bundles containing medium-sized to very fine nerve fibers that become finer and narrower toward the mu-cosa. Aggregations of neurons form small intramural ganglia at the intersection of fibers.
The intramural ganglia (E) contain primarily multipolar, rarely unipolar, neurons with granular Nissl substance. The neurons are surrounded by flat sheath cells, and their numerous long dendrites are thin and often indistinguishable from axons. The axons are extremely thin, unmyelinated or poorly my-elinated, and often arise from a dendrite rather than the perikaryon. Between the neurons lies a dense network of fibers in which it is difficult to differentiate between dendrites, terminating axons, and fibers passing through the ganglion. Sympathetic and parasympathetic neurons have the same shape; they can only be distinguished histochemically.
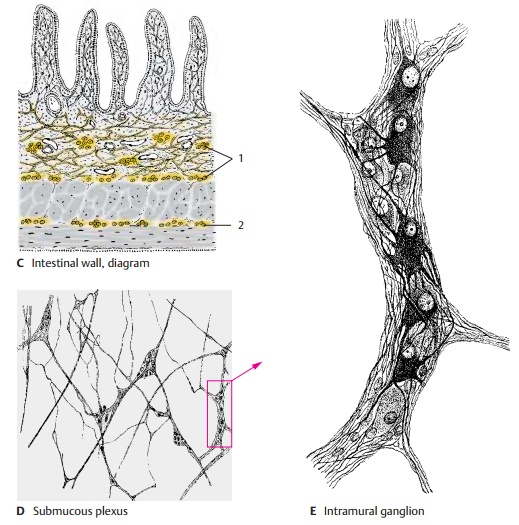
The myenteric plexus (C2) is embedded in a narrow space between the transverse and longitudinal muscles of the intestine. It con-sists of a relatively regular network of coarse and fine fiber bundles. Apart from the intramural ganglia at the intersections, there are numerous neurons that are often arranged in rows along the fiber bundles.
The enormous number of neurons scattered throughout the tissue almost amounts to an independent nervous organ. This explains the local autonomy of the gastrointestinal tract, which remains functional even after denervation.
As derivatives of the neural crest, the paraganglia and the adrenal medulla are regarded as part of the autonomic nervous system (see vol. 2).
Autonomic Neurons
Structure (A, B, D)
The autonomic nervous system consists of numerous individual elements, the auto-nomic neurons (A) (neuron doctrine). The theory of neuronal continuity, which hadlong been postulated for the terminal rami-fications of the autonomic nervous system, has been disproved by electron-microscopic studies. According to the continuity theory, the terminal ramifications of an intramural plexus were thought to form a continuous network (terminal reticulum), in which the processes of different neurons would fuse with each other and with the innervated muscle cells and glandular cells. The net-work was thought to represent a syncytium where nerve fibers share a common cyto-plasm. The electron-microscopic image does not show such continuity.
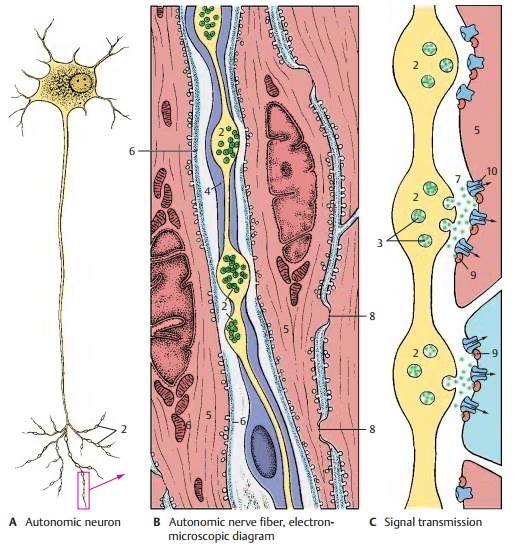
However, the postganglionic neurons do have specific features. The bundles of nerve fibers exhibit numerous axo-axonal syn-apses (D1), not only among sympathetic andparasympathetic fibers themselves but also between sympathetic and parasympathetic fibers. In the area of terminal ramification, specific structures corresponding to the motor end plates at striated muscles are absent. The only remarkable features are varicose swellings (varicosities) (A – D2) along the terminal branches of the axons.
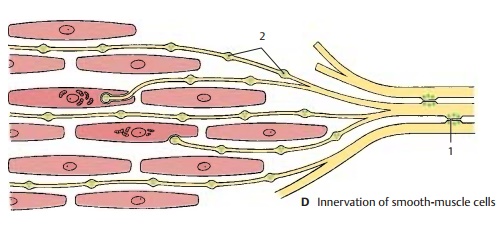
The axon swellings may lead to indenta-tions on the smooth-muscle cells or may even invaginate the cells. In general, however, they lie between the muscle cells without direct membrane contact (as they would in a synapse, ). The swell-ings contain clear and granular vesicles (C3) similar to those in presynaptic terminal boutons The granular vesicles have been shown to contain norepinephrine, the neurotransmitter of the sympathetic nervous system. The sheath of Schwann cells(B4) surrounding the terminal branches is absent in the area of the swelling, and the adjacent wall segment of the smooth-muscle cell (BC5) lacks a basement mem-brane (B6). This is the site of signal trans-mission from the autonomic nerve fiber to the smooth-muscle cell.
Signal Transmission (C)
The vesicles contained in the axon swellings release their content into the intercellular space (C7). The neurotransmitter molecules diffuse into the intercellular space, thereby transmitting the signal to a large number of smooth-muscle cells. Signal transmission also spreads via membrane contacts be-tween the muscle cells. The smooth-muscle cells are interconnected by gap junctions (B8) which function as electrical synapses (see p. 26). There is no basement membrane at gap junctions.
Efferent autonomic nerve fibers innervate the smooth muscles and glandular cells (secretory fibers). Innervation of glandular cells (blue cells in C) is essentially the same as that of muscle cells. The neurotransmit-ters released from the vesicles in the axon swellings (green dots in C) activate G pro-tein-coupled receptors (C9) on the surface of the innervated cell. G protein mediates the opening of ion channels (C10), thus trig-gering an intracellular cascade of signals. This results in the contraction of smooth-muscle cells or in the synthesis and release of glandular secretion by the glandular cells
Related Topics