Chapter: Medical Microbiology: An Introduction to Infectious Diseases: Viral Structure
Virion Size and Design - Viral Structure
VIRION SIZE AND DESIGN
Viruses are approximately 100- to 1000-fold smaller than the cells they infect. The small-est viruses (parvoviruses) are approximately 20 nm in diameter (1 nm 10 -9 m), whereas the largest animal viruses (poxviruses) have a diameter of approximately 300 nm and overlap the size of the smallest bacterial cells (Chlamydia and Mycoplasma).
There-fore, viruses generally pass through filters designed to trap bacteria, and this property can, in principle, be used as evidence of a viral etiology.
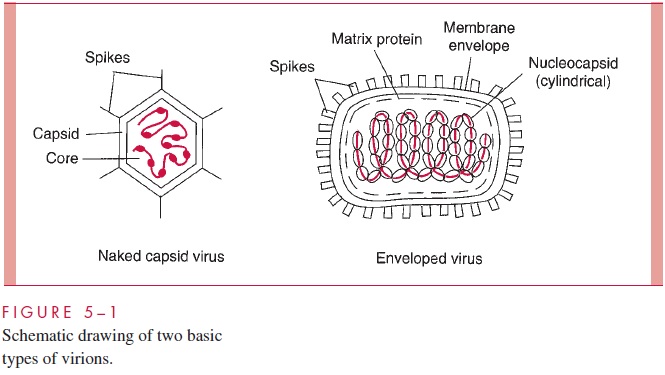
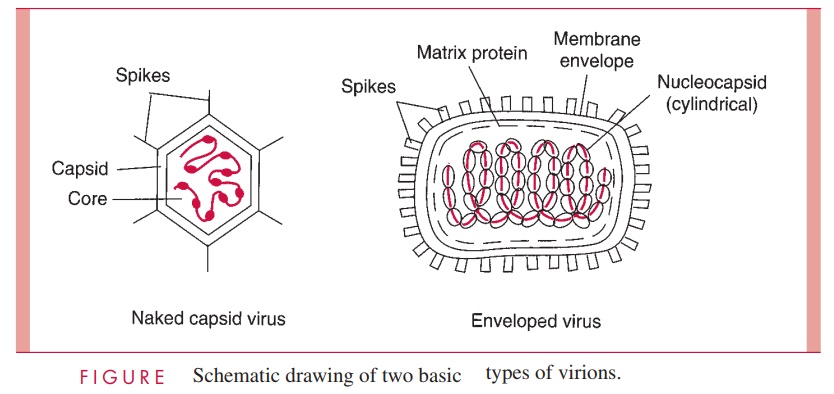
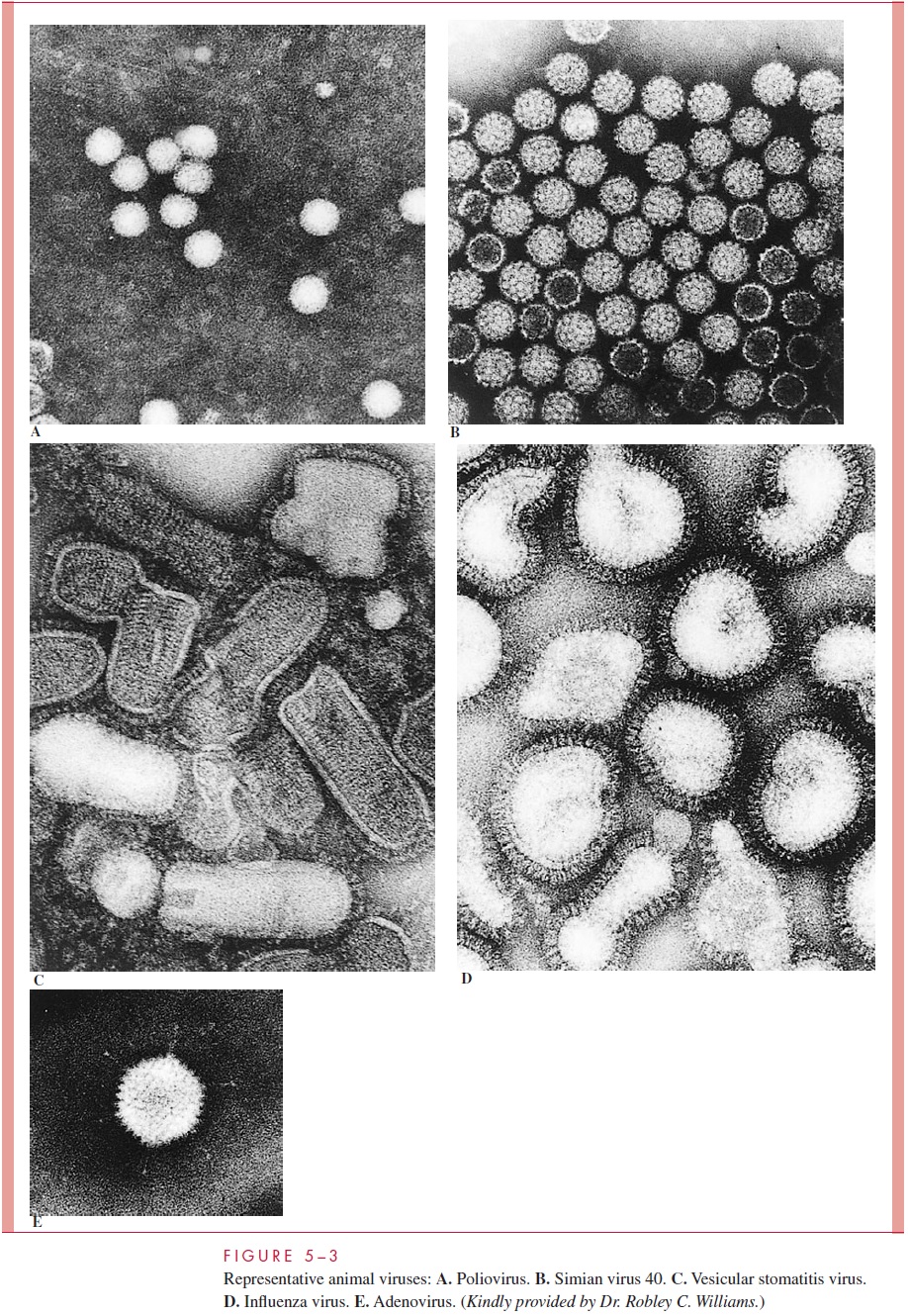
The basic design of all viruses places the nucleic acid genome on the inside of a protein shell called a capsid. Some animal viruses are further packaged into a lipid mem-brane, or envelope, which is usually acquired from the cytoplasmic membrane of the infected cell during egress from the cell. Viruses that are not enveloped have a defined external capsid and are referred to as naked capsid viruses.The genomes of enveloped viruses form a protein complex and a structure called a nucleocapsid, which is often sur-rounded by a matrix protein that serves as a bridge between the nucleocapsid and the inside of the viral membrane. Protein or glycoprotein structures called spikes, which often protrude from the surface of virus particles, are involved in the initial contact with cells. These basic design features are illustrated schematically in Figure 5 – 1 as well as in the electron micrographs in Figures 5 – 2 and 5 – 3.
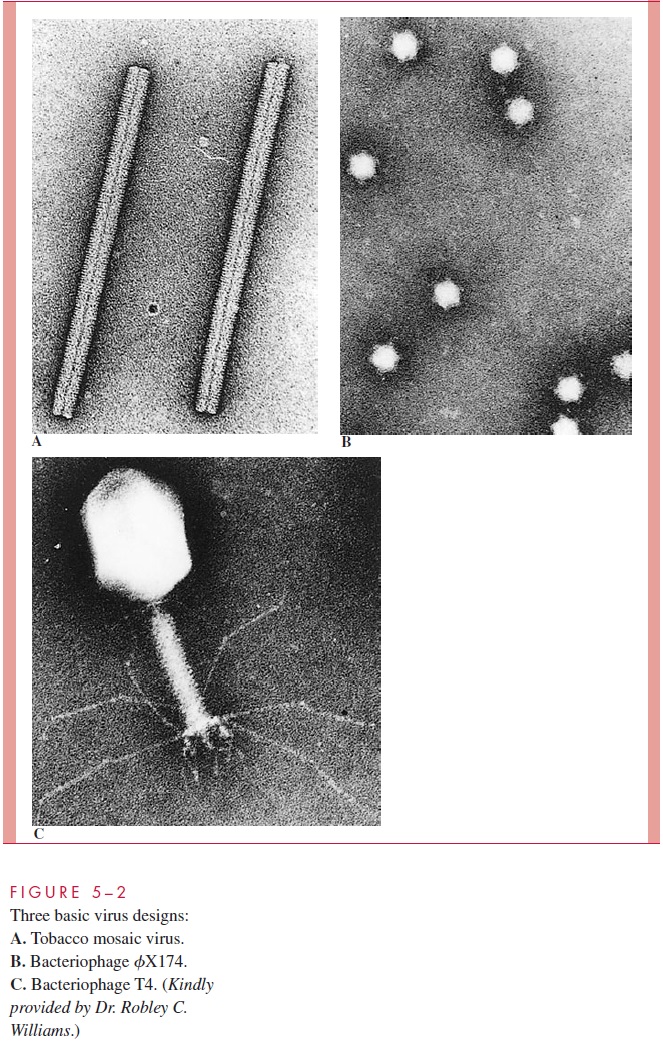
The protein shell forming the capsid or the nucleocapsid assumes one of two basic shapes: cylindrical or spherical. Some of the more complex bacteriophages combine these two basic shapes. Examples of these three structural categories can be seen in the electron micrographs in Figure 5 – 2.
The capsid or envelope of viruses functions (1) to protect the nucleic acid genome from damage during the extracellular passage of the virus from one cell to another, (2) to aid in the process of entry into the cell, and (3) in some cases to package enzymes essen-tial for the early steps of the infection process.
In general, the nucleic acid genome of a virus is hundreds of times longer than the longest dimension of the complete virion. It follows that the viral genome must be exten-sively condensed during the process of virion assembly. For naked capsid viruses, this condensation is achieved by the association of the nucleic acid with basic proteins to form what is called the core of the virus (see Fig 5 – 1). The core proteins are usually en-coded by the virus, but in the case of some DNA-containing animal viruses, the basic pro-teins are histones scavenged from the host cell. For enveloped viruses, the formation of the nucleocapsid serves to condense the nucleic acid genome.
Two classes of infectious agents exist that are structurally simpler than viruses. Viroids are infectious circular RNA molecules that lack protein shells; they are re-sponsible for a variety of plant diseases. Hepatitis delta, an infectious agent sometimes found in association with hepatitis B virus, appears to share many properties with the viroids. Prions, which apparently lack any genes and are composed only of protein, are agents that appear to be responsible for some transmissible and inherited spongi-form encephalopathies such as scrapie in sheep; bovine spongiform encephalopathy in cattle; and kuru, Creutzfeldt-Jakob disease, and Gerstmann-Sträussler-Scheinker syn-drome in humans.
Related Topics