Chapter: Medical Microbiology: An Introduction to Infectious Diseases: Spirochetes
Spirochetes - Bacteriology
BACTERIOLOGY
MORPHOLOGY AND STRUCTURE
The spiral morphology of spirochetes is produced by a flexible, peptidoglycan cell wall around which several axial fibrils are wound. These fibrils have the structure of flagella and are referred to as endoflagella (Fig 27 – 1). The cell wall and endoflagella are com-pletely covered by an outer bilayered membrane similar to the outer membrane of other Gram-negative bacteria. In some species, a hyaluronic acid slime layer forms around the exterior of the organism and may contribute to its virulence. Spirochetes are motile, exhibiting rotation and flexion; this motility is believed to result from movement of the endoflagellar filaments, although the mechanism is not clear.
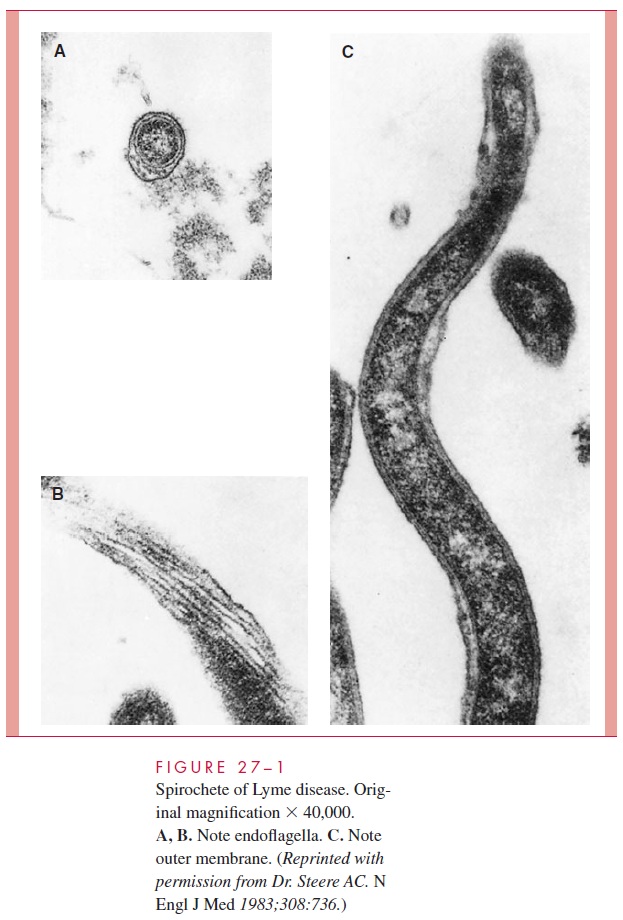
Many spirochetes are difficult to see by routine microscopy. Although they are Gram-negative, many either take stains poorly or are too thin (0.15μm or less) to fall within the resolving power of the light microscope. Only darkfield microscopy, immunofluorescence, or special staining techniques that effectively increase their diameter can demonstrate these spirochetes. Other spirochetes such as Borrelia are larger and readily visible in stained preparations, even routine blood smears.
GROWTH AND CLASSIFICATION
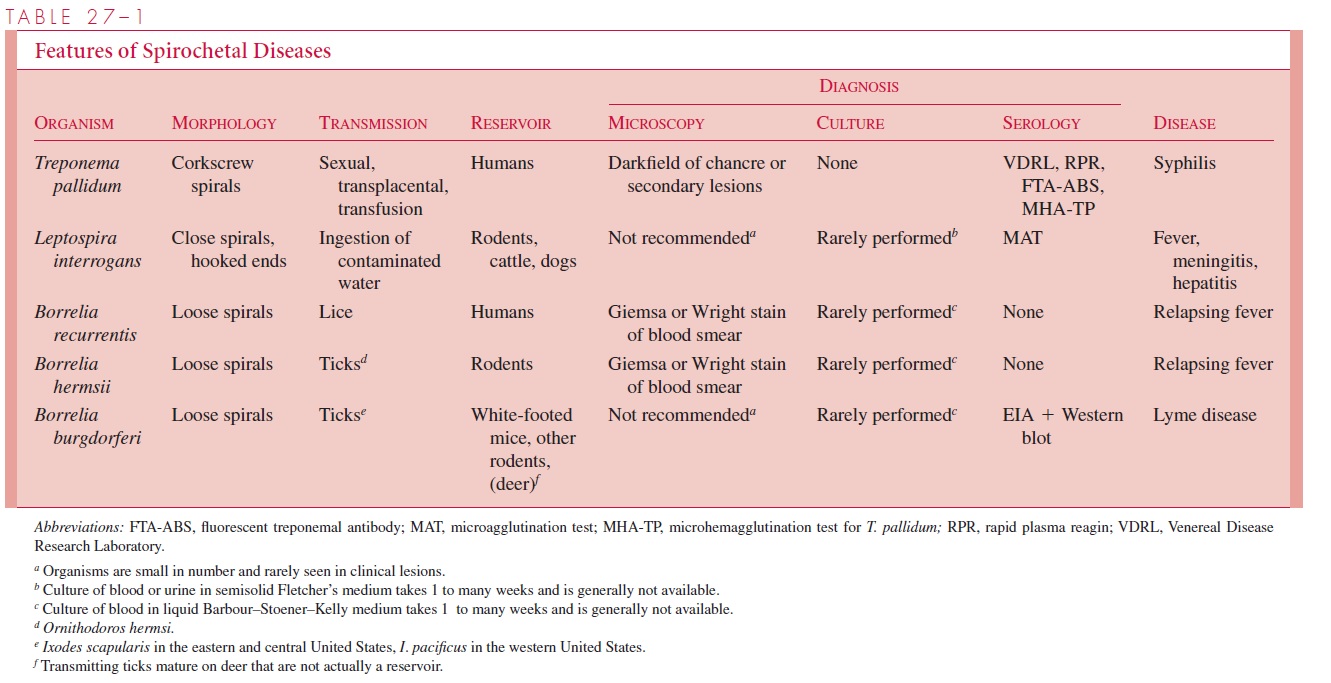
Parasitic spirochetes grow more slowly in vitro than most other disease-causing bacteria. Some species, including the causative agent of syphilis, have not been grown beyond a few generations in cell culture. Some are strict anaerobes, others require low concentrations of oxygen, and still others are aerobic. Compared to other bacterial groups the taxonomy of the spirochetes is underdeveloped. Because spirochetes are difficult to grow, they are diffi-cult to study; thus, there are relatively few phenotypic properties on which to base a classi-fication. The medically important genera Treponema, Leptospira, and Borrelia have been distinguished primarily by morphologic characters such as the nature of their spiral shape and the arrangement of flagella. Modern DNA homology and ribosomal RNA analyses have supported these groupings.
Related Topics