Chapter: Human Neuroanatomy(Fundamental and Clinical): Sensory Receptors and Neuromuscular Junctions
Proprioceptive Receptors
Proprioceptive Receptors
Golgi Tendon Organs
They are also called the neurotendinous organs of Golgi. These organs are located at the junction of muscle and tendon. Each organ is about 500 μm long and about 100 μm in diameter. It consists of a capsule made up of concentric sheets of cytoplasm (Fig. 4.2). Inside the capsule there are small bundles of tendon fibres. The organ is innervated by one or more myelinated nerve fibres that divide to form several branches (spray-like arrangement). These receptors are stimulated by pull upon the tendon during active contraction of the muscle, and to a lesser degree by passive stretching.
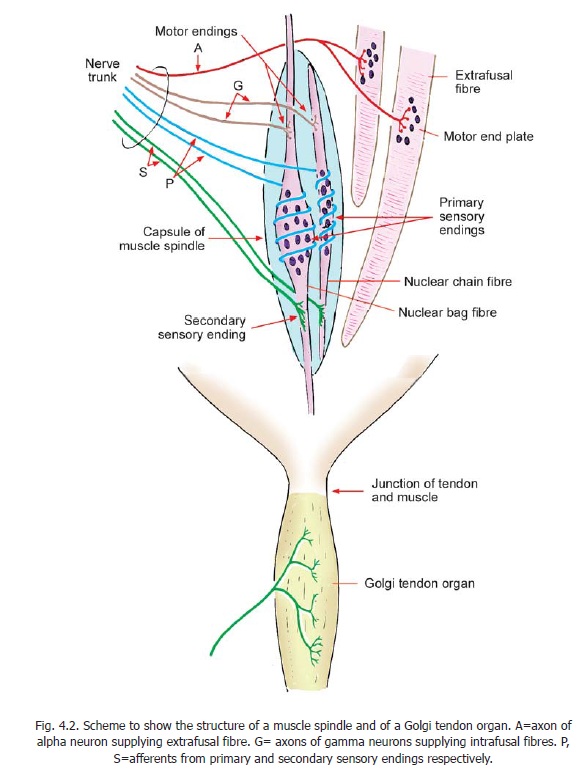
In the past Golgi tendon organs have been considered to be involved in myotactic reflexes that prevent the development of excessive tension in muscle. However, it is now believed that their role is mainly in providing proprioceptive information; and that they are slow adapting receptors.
Similar endings are also present in ligaments of joints. At this site they serve as slow adapting, high threshold, receptors. Impulses from them lead to reflex inhibition of adjacent muscles, preventing excessive stresses on ligaments.
Muscle Spindles
These are spindle-shaped sensory end organs located within striated muscle (Fig.4.2). The spindle is bounded by a fusiform connective tissue capsule (forming an external capsule) within which there are a few muscle fibres of a special kind. These are called intrafusal fibres in contrast to extrafusal fibres that constitute the main bulk of the muscle. Each spindle contains six to fourteen intrafusal fibres. Each intrafusal fibre is surrounded by an internal capsule of flattened fibroblasts and collagen.
Intrafusal fibres contain several nuclei that are located near the middle of the fibre. In some fibres this region is dilated into a bag: these are nuclear bag fibres. In other intrafusal fibres the nuclei lie in a single row, there being no dilatation: these are nuclear chain fibres.
Each muscle spindle is innervated by sensory as well as motor nerves. The sensory endings are of two types, primary and secondary. The motor innervation of intrafusal fibres is (mainly) by axons of gamma neurons located in the ventral grey column of the spinal cord . The sensory endings respond to stretch. Primary sensory endings are rapidly adapting while secondary endings are slow adapting. However, the precise role of these receptors is complex and varies in different types of fibres.
Spindles provide information to the CNS about the extent and rate of changes in length of muscle. Nuclear bag fibres are stimulated by rapid changes, while nuclear chain fibres react more slowly. Contraction of intrafusal fibres makes the spindle more sensitive to stretch.
Advanced
Some further details about muscle spindles are as follows.
1. The nuclear bag fibres are considerably larger than the nuclear chain fibres. They extend beyond the capsule and gain attachment to the endomysium of extrafusal fibres. The nuclear chain fibres, on the other hand, remain within the capsule to which their ends are attached. On the basis of their ultrastructure and physiological properties nuclear bag fibres are divided into two types. Bag1 (or dynamic bag1) fibres respond to rapid changes in muscle length. Bag2 (or static bag2) fibres are less responsive to such changes.The various types of intrafusal fibres differ from extrafusal fibres, and from each other, in ultrastructure and in some features of histochemistry.
2. The primary sensory fibres wind spirally around the nuclear region of intrafusal fibres and are, therefore, referred to asannulospiral endings. The secondary endings (also called flower sprayendings) are seen mostly on nuclear chain fibres and are located away from the nuclear region. Bothprimary and secondary nerve fibres are derived from large myelinated axons, but are themselves unmyelinated.
3. The motor endings on intrafusal fibres of muscle spindles are of three types.
a. Terminals of gamma-efferents that end on the equator of the nuclear bag, and do not show typical end plates.
b. Gamma-efferents ending some distance away from the equator of the nuclear bag and having typical end plates. These are also called P2 endings.
c. Terminals of delta-efferents (equivalent to beta-efferents of some species), which are collaterals of alpha-fibres supplying extrafusal muscle fibres. These terminals are located near the ends of nuclear bag fibres. They are also called P1 endings.
4. The motor nerve fibres innervating intrafusal fibres are thin but are myelinated. Those ending over nuclear bags do not show end plates. The P2 endings show typical end plates. P1 endings show en grappe end plates.
Receptors present in relation to joints
Four types of receptors have been demonstrated in relation to joints.
1. Type I. These resemble Ruffini endings. They are innervated by myelinated nerve fibres, andserve as slowly adapting mechanoreceptors. These receptors are responsible for the sense of joint position and movement.
2. Type II. These are similar to Pacinian corpuscles. They are fast adapting mechano-receptors,supplied by myelinated nerve fibres.
3. Type III. These are similar to neurotendinous organs of Golgi. Impulses arising in them areprobably responsible for reflex inhibition of muscle contraction, thus preventing excessive movement.
4. Type IV. These are free nerve endings, probably responsible for pain.
Related Topics