Chapter: Anatomy of Flowering Plants: An Introduction to Structure and Development : Organs, Cells and Tissues
Plant Vascular Tissue: xylem and phloem
Vascular Tissue
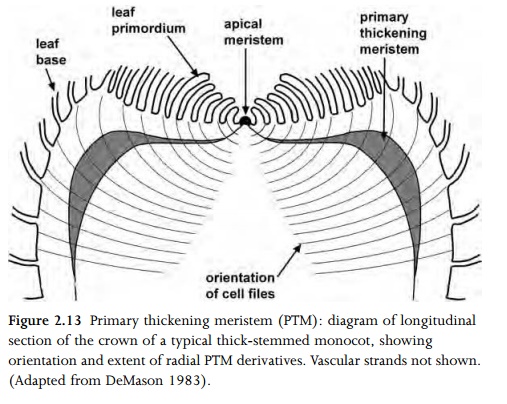
Vascular tissue consists of xylem and phloem, and may be primary or secondary in origin. Primary vascular tissue is derived from procambium, itself produced by the apical meristems, and also by the primary thickening meristem in stems of monocots. Secondary vascular tissue is derived from the vascular cambium in dicots, and from the secondary thickening meristem in a few monocots (Fig. 2.13). Both xylem and phloem are complex tissues, composed of many different cell types. Xylem is primarily concerned with water transport and phloem with food transport. Distribution of vascular tissue varies considerably between different organs and taxa.
Xylem
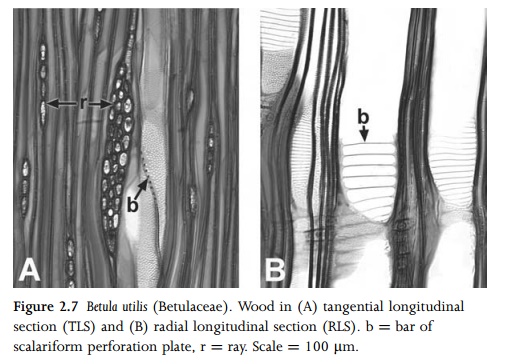

These pits vary considerably in size, shape and arrangement; they may be oval, polygonal or elongated (scalariform pitting), organized in transverse rows (opposite pitting) or in a tightly packed arrangement (alternate pitting).
Phloem
Phloem has complex roles in translocation and messaging within the plant. Primary phloem is formed by the apical meristem and secondary phloem by the vascular cambium. Phloem may develop precociously in regions that require a copious supply of nutrients, such as developing sporogenous tissue.
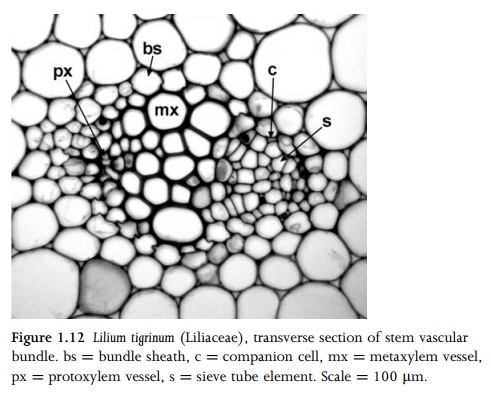
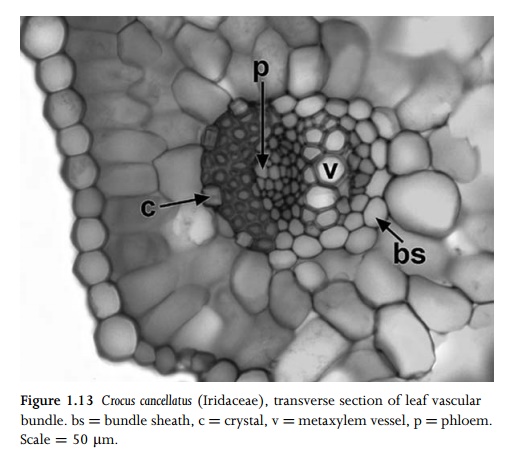
Phloem is a complex tissue that consists of conducting cells (sieve elements) and associated specialized parenchyma cells (companion cells) (Figs. 1.12; 1.13); these two closely inter-dependent cell types are produced from a common parent cell but develop differently. Angiosperm sieve elements lack nuclei and most organelles at maturity, but retain plastids and phloem-specific
proteins (P-proteins) which occur in several morphological forms (amorphous, filamentous, tubular and crystalline) that are often highly characteristic for particular plant families, and thus of systematic and evolutionary value. Sieve-element plastids are classified according to their inclusions: starch (S-type plastids), protein (P-type plastids), or both. By contrast, companion cells are densely cytoplasmic, retaining nuclei and many active mitochondria.
Sieve elements are linked axially to form sieve tubes. The two basic types of sieve element, sieve cells and sieve-tube elements, are differentiated by their pore structure; most angiosperms exclusively possess sieve-tube elements. The walls of sieve ele-ments are thin and possess characteristic regions (sieve areas) that connect adjacent sieve elements; sieve areas consist of groups of pores and associated callose. In sieve cells the sieve areas are distributed throughout the cell wall, but in sieve-tube elements they are mainly localized on the adjoining end walls, forming sieve plates that link two axially linked elements of a sieve vessel. Sieve plates can be simple or compound.
Related Topics