Chapter: Plant Biology : Plant communities and populations
Plant Populations
POPULATIONS
Key Notes
Plant life cycles
Plants may live for a few weeks to thousands of years; reproduce once and die or reproduce many times. Semelparous plants reproduce just once, after a few weeks (ephemerals) or over a year (biennials) to many years (semelparous perennials). Iteroparous plants reproduce many times and some of these produce rhizomes and spread clonally.
Ecology of ephemeral plants
These occur everywhere but are common in temperate climates and deserts. Many have seeds that can remain dormant for many years and their numbers often fluctuate markedly between years. Many temperate species have small self-fertilizing flowers.
Biennials and semelparous perennials
Biennials are opportunist plants, often with seeds that can be dormant, living in successional habitats mainly in the temperate zone. They produce showy flowers. Life cycles are flexible and they live longer if damaged. Semelparous perennials live in conditions where growth is slow or where swamping seed predators is important.
Iteroparous Perennials
These range from short-lived herbaceous species similar to ephemerals to long-lived trees. They dominate most stable habitats and, in any one place, the dominants often have individuals all of similar age owing to cyclical changes in the plant community. Some spread clonally, particularly herbaceous plants on woodland floors and in wetlands
Timing of Reproduction
Early reproduction is often an advantage but iteroparous perennials are at an advantage if there is high seedling mortality.
Population dynamics
There are many hazards in a plant’s life cycle, but all populations have the potential to increase exponentially, the rates depending on birth rate, death rate, immigration and emigration. If plants grow above a certain density, over time density-dependent mortality occurs, known as selfthinning. This can be described mathematically with the –3/2 power law which is constant for many plants. There is often great variation in size within a population and small plants die first.
Populations of clonal Plants
A ‘population’ of a clonal plant can be one genetic individual. Clonal plants can regulate their own shoot density although sometimes shoots compete. Growth behavior differs in different environments and they can exploit rich patches effectively while crossing less favorable ones. The disadvantage of clonal growth is mainly disease susceptibility.
Plant life cycles
A complete life cycle of a plant, from germination through its reproductive life to death, varies from about 3 weeks to several thousand years. Plants may be divided into those that flower and set seed once in their lives, known as semelparous or monocarpic, and those that reproduce more than once, known as iteroparous or polycarpic. Semelparous plants may live for a few weeks or for many years. Short-lived ones are known asephemerals or, commonly, annuals, although most complete their life cycle in much less than a year, and the shortest-lived, such as Boerhavia spp. (Nyctaginaceae) may complete their lifecycle in less than 4 weeks. Those that live longer than a year are mostly known as biennials. These normally grow as a leaf rosette in their first year, flowering and dying in their second. Longer-lived semelparous plants are rarer but some of these have leaf rosettes of ever increasing size over several years, such as the century plants, Agave spp. (Agavaceae), which usually live for 15–70 years rather than a century. Some are woody, notably some bamboos which can flower after 120 years, and the royal palm, Roystonea, which can live for 80 years and reach 30 m before flowering and dying. Those that live for more than about 3 years are known as semelparous perennials.
Iteroparous plants range from those that live for just a few years and normally flower less than five times to some of the longest-lived of all organisms such as the huon pine, Dacrydium franklinii, in Tasmania and the bristlecone pine, Pinus aristata, in Arizona, both of which can live for over 4000 years. Living creosote bushes, Larrea tridentata (Zygophyllaceae), of the deserts of western USA may be even older, over 11 000-years-old, spreading clonally from a central source. Nearly all woody plants are iteroparous.
All plants have meristems and if shoots remain vegetative they have indeterminate growth, i.e. continue to grow throughout their lives, remaining ‘forever young’. Many have developed underground rhizomes or stolons which grow continuously and can form large patches or grow a long way from where they first germinated. These produce roots as they grow and the plant can then be split to form independent physiological units. These clones, or parts of them, can live for many centuries and the definition of an individual becomes obscured since a genetic individual and a physiological individual become different. Clonal spread is frequent among herbaceous perennials, less so among woody plants.
Ecology of ephemeral plants
These occur in most plant communities but are common in deserts, regions with a Mediterranean climate of cool wet winters and hot dry summers, agricultural and disturbed land and places with unstable soils like sand dunes. In mature plant communities such as woodlands and permanent grasslands they are mainly associated with disturbed areas, e.g. from animal digging or tree falls. Some will germinate at almost any time of year and desert ephemerals germinate in response to a substantial fall of rain. There is a group of ‘winter annuals’ in temperate climates that germinate in the fall and flower and set seed in the spring before the summer drought. In many ephemerals, generations of adult plants do not overlap, but they may have seeds that can remain dormant for a long period so seeds from different years may germinate together. Some appear to rely on each year’s seed crop and have limited dormancy. Many ephemerals in temperate zones are small plants with small flowers that can self-fertilize, but among desert ephemerals there is great variation and many have large colorful flowers. Most populations of ephemeral plants fluctuate markedly from year to year.
Biennials and semelparous perennials
Biennials are opportunist plants, like the ephemerals, and many have long-lived dormant seeds. They occur in early successional stages where a mature habitat such as a woodland or grassland has been disturbed from tree falls for example, or at the edges of grasslands, mainly in temperate climates. They do not occur on agricultural land in general. Frequently the life cycle is flexible and a normally biennial plant can perennate (i.e. live for longer and flower a second time) if its flowers or fruits are eaten or damaged. Frequently the flowers are produced in showy inflorescences attractive to flower-visiting animals, this being especially true for those plants that live for longer than a year before flowering. By the time they flower, the place they are occupying is often not suitable for the next generation so the seeds must disperse or lie dormant.
Some semelparous perennials are associated with regions where growth is restricted such as deserts or tropical mountains; others such as the bamboos are thought to reproduce once so that the huge numbers of seeds produced will swamp any potential predators. Frequently in these species all the plants in an area will flower synchronously.
Iteroparous perennials
These range from short-lived pioneers that behave in a similar way to ephemeral and biennial species through to long-lived species with much more stable populations. The dominant species in most communities are long-lived non-clonal species, such as most trees and many of the dominant plants of open areas. The adult plants of the dominant species in some communities are of similar age where there is some kind of cyclical change and the main germination is at a time when the area is more open, through a natural disaster. Understorey trees and other plants in a woodland, by contrast, normally have a wide range of ages including many seedlings and saplings often with smaller numbers of adults.
Clonal species dominate certain habitats, particularly wetlands and woodland floor habitats, and one genetic individual can cover a large area, even an entire habitat. The vegetative branches, or ramets, can utilize the resources of the mother plant as they grow and this gives them a huge advantage in establishing themselves over non-clonal plants that rely on new seedlings.
Timing of reproduction
In general, it will be selectively advantageous for a plant to reproduce as early as possible. This will often lead to maximal population growth, e.g. if a plant produces 10 seeds after 1 year and then dies, the following year each of those 10 seeds can grow and produce 10 seeds, so any plant that delays reproduction until the second year will need to produce 100 seeds to equal the potential output of one reproducing in its first year. Although this does not take into account mortality at any stage, which is likely to be different for seeds and established plants, the potential advantage of reproducing early is clear and ephemerals are thought to be among the most advanced of plants. Ephemeral plants put most of their resources into reproduction, but an iteroparous perennial needs to retain some resources for its own survival so seed production is likely to be less. To determine which is at an advantage the survival of seedlings (pre-reproductive) and of adults must be taken into account. We can say that for an ephemeral plant:
le = cme
Where le is the rate of increase, c is the seedling survival and me is the mean seed production.
For an iteroparous perennial:
lp = cmp + p
where p is the adult survival.
We can derive from this that an ephemeral plant will reproduce faster if me >mp + p/c. This indicates that one of the important terms is the ratio between adult survival and seedling survival (p/c): if there is good seedling survival it will be advantageous to be ephemeral; if seedling survival is poor an iteroparous perennial will be at an advantage. Studies have confirmed that this is largely true.
Population dynamics
There is often high mortality of the seeds, and there may be many natural hazards before they can germinate, such as seed-eating animals, rain and wind. After germination, the main competition occurs during active growth to flowering, and this is likely to involve competition with associated plants as well as members of the same species. After this an established plant faces many fewer hazards, so the greatest selection is likely to be in the seedling stage. In general, plants live in populations in the same way as animals, with birth rates, death rates, immigration and emigration. Any change in that population must take these into account and this can be expressed by this basic equation of population change:
Nt+1 = Nt + B – D + I – E
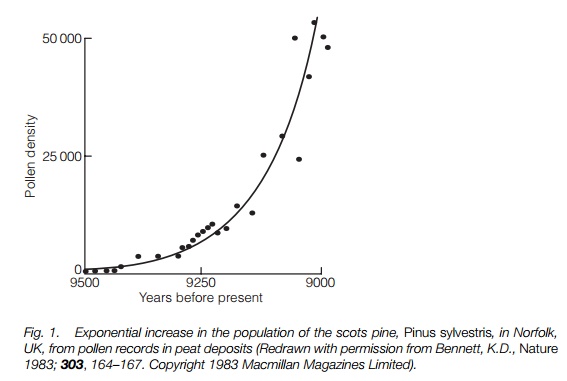
where Nt is the number at time t, B is number of births, D number of deaths, I is immigration, E is emigration. When B + I = D + E the population is stable, but when B + I >D + E the population will increase exponentially, i.e. increasing by the same factor each year, giving a logarithmic growth curve (Fig. 1). This has been recorded when a plant is colonizing a new area. Clearly this rate of increase can only continue for a short period after which the population will be regulated by some finite resource such as germination sites or nutrient availability.
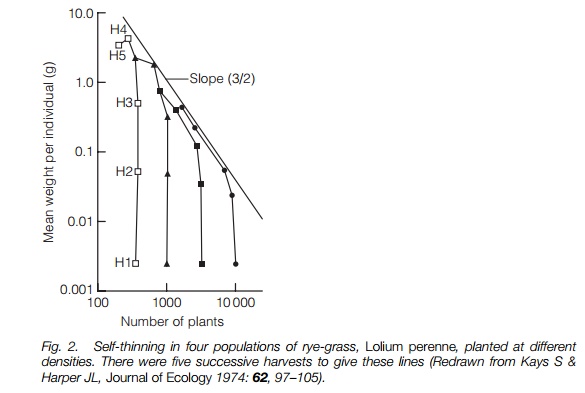
As density increases the plants may start to interact with each other, limiting each others growth. This may be manifest in smaller plants and/or lower seed set, particularly in small ephemerals, but with longer lived plants some will die before reaching maturity. This is most obviously seen in trees where seedlings can cover the ground but there is fewer than one mature tree per 10 m2. There is a process of self-thinning. The idea has been best developed in relation to crop plants and their yields since farmers will want to sow a crop to reach maximal yield while minimizing seed wastage. Experimentally, it was found that mortality started earlier, and at a smaller weight with denser sowings. If the relationship between sowing density and mean weight is plotted using a log scale on both axes the lines converge on a straight line (Fig. 2). This line has a slope of approximately –3/2 for many unrelated plants (including trees) and can be described by the equation:
log w = c – 3/2 logN
where w is mean plant weight, c is a constant, N is plant numbers. This is known as the –3/2 power law. In sparse populations interaction between individuals will be less and the slope of the line will flatten out, until it reaches 0 when there is no density-dependent interaction. Although this slope is remarkably constant across plants, the c value varies hugely. This is the intercept term on the y axis, i.e. the density at which there is no interaction between individuals, clearly totally different for a small ephemeral and a tree. There are other factors that influence numbers of plants that are dependent on the density of the population, such as disease and herbivore attack. Their effects are often intermittent and they are much harder to study than competition, but they can be equally or more important in determining population density .
If a population of a large plant such as a tree starts out at high density, over time there will arise a marked size difference between the largest and the smallest individuals, and the smallest will be the first to die in the self-thinning
process unless some other influence intervenes. Initial size difference may arise from slightly different germination times or genetic differences between individuals in their vigor of growth but this will be accentuated by competition once they start interacting.
In general, a plant population is affected by many processes that are independent of a plant’s density such as frosts or floods at particular times and indensity-dependent interactions with members of its own and other species.
Populations of clonal plants
Plants that spread vegetatively can regulate the density of their own shoots, although there can be competition between shoots as described for non-clonal species. An entire ‘population’ can be made up of just one genetic individual and this may be true of a hillside covered with bracken, Pteridium aquilinum, or a reed bed dominated by Phragmites australis. Clonal plants may grow densely to dominate an area or as long strands invading new places, and these invaders can tap the resources of the mother plant giving them a great advantage over seedling invaders. The invaders can change their ‘behavior’ (in a plant, behavior means changes in growth form) in response to local conditions, growing densely where there are many nutrients, but sparsely between rich patches.
There appears superficially to be such a great advantage to clonal spread that it is surprising that, although it is common, more plants do not spread this way. The main disadvantage lies in the fact that clones are all identical genetically. This leaves them susceptible to insect attack or disease since, if a predator or pathogen can overcome the resistance of one member of the clone, all will be susceptible. This was shown graphically in Europe in the 1970s with the outbreak of elm disease attacking all the clonal elm species, e.g. Ulmus procera, but only some individuals of the mainly non-clonalUlmus glabra. In agriculture, clonally produced crops require greater uses of pesticides.
Related Topics